与无Tat标签蛋白相比,Tat标签蛋白的α-螺旋和转角显著增加,β-折叠和无规卷曲含量显著降低
为了阐明SodA、SodB和SodC与Tat标签结合时的二级结构变化,通过Ni-NTA亲和层析纯化了Tat标签蛋白和无Tat标签蛋白。色谱图和洗脱蛋白如图所示。制备了高度富集的纯化蛋白样品,并通过圆二色性测定了Tat标签蛋白和无Tat标签蛋白的二级结构。如直方图结果所示,Tat标签蛋白的二级结构与无Tat标签蛋白不同:与无Tat标签蛋白相比,Tat标签蛋白的α-螺旋和转角显著增加,而β-折叠和无规卷曲含量显著降低。
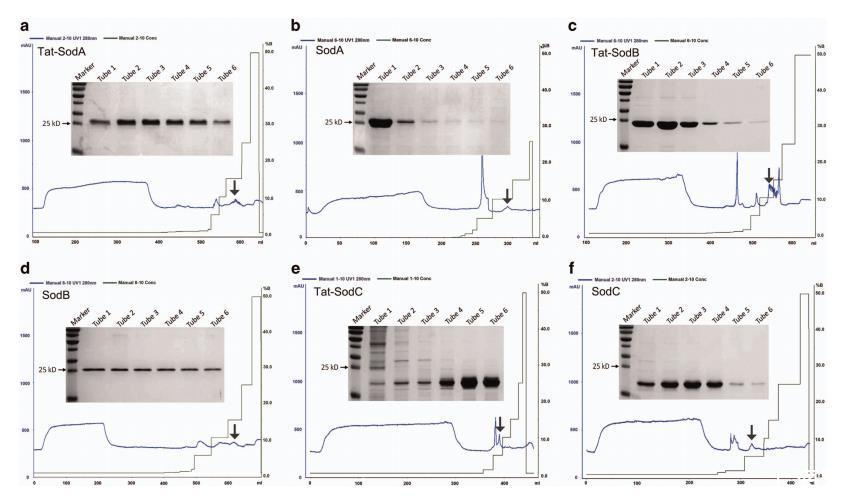
图 7 色谱图及通过聚丙烯酰胺凝胶电泳鉴定的洗脱蛋白。(a) Tat-SodA的色谱图及通过SDS-PAGE鉴定的洗脱蛋白。(b) SodA的色谱图及通过SDS-PAGE鉴定的洗脱蛋白。(c) Tat-SodB的色谱图及通过SDS-PAGE鉴定的洗脱蛋白。(d) SodB的色谱图及通过SDS-PAGE鉴定的洗脱蛋白。(e) Tat-SodC的色谱图及通过SDS-PAGE鉴定的洗脱蛋白。(f) SodC的色谱图及通过SDS-PAGE鉴定的洗脱蛋白。
在我们的研究中,我们证明了添加Tat标签促进了大肠杆菌中融合蛋白的高产量和可溶性表达,并且Tat标签蛋白的总SOD活性显著高于无Tat标签蛋白。此外,过表达Tat标签SOD蛋白的细菌细胞的抗百草枯活性显著高于表达无Tat标签蛋白的细胞,并且ΔSodA突变菌株的生长在百草枯处理后受到显著抑制。与编码无Tat标签蛋白的基因相比,编码Tat标签蛋白的基因的mRNA转录水平显著增加。此外,与无Tat标签蛋白相比,Tat标签蛋白的α-螺旋和转角含量显著增加,而β-折叠和无规卷曲含量显著降低。这些结果表明,添加Tat标签可以增加mRNA转录水平,促进异源蛋白在大肠杆菌中的高产量和可溶性表达,并且可以改变其融合蛋白的二级结构。
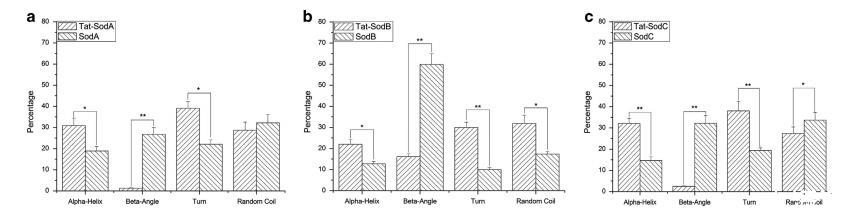
图 8 通过圆二色性对Tat标签蛋白和无Tat标签蛋白进行二级结构测定。(a) 通过CD对Tat-SodA和SodA进行二级结构测定。(b) 通过CD对Tat-SodB和SodB进行二级结构测定。(c) 通过CD对Tat-SodC和SodC进行二级结构测定。星号表示蛋白质间的α-螺旋、转角、β-折叠和无规卷曲含量具有统计学显著差异 (*P<0.05; **P<0.01)。
HIV-1编码的Tat蛋白是蛋白质转导结构域家族的成员,富含碱性氨基酸。众所周知,全长Tat肽和核心结构域在异源生物大分子如蛋白质、肽和核苷酸跨体内各类生物膜的转导中起重要作用。Tat的核心区域包含一个含有六个精氨酸和两个赖氨酸的碱性氨基酸结构域。目前的研究表明,异源蛋白的跨膜转导效率立即受到氨基酸数量的影响。蛋白质二级结构预测结果显示,在Tat碱性氨基酸结构域中可以形成α-螺旋结构。先前的研究表明,Tat蛋白转导肽的碱性氨基酸结构域不能形成功能结构。二级结构可以通过在原子分辨率结构中观察到的蛋白质氢键模式来正式定义。α-螺旋在DNA结合基序中具有特殊意义。β-折叠是最常见的蛋白质结构元件,对正确折叠至关重要。在我们的研究中,圆二色性结果表明,与无Tat标签蛋白相比,Tat标签蛋白的α-螺旋和转角比例显著增加,而β-折叠和无规卷曲含量显著降低,表明添加Tat标签可以改变其融合蛋白的二级结构。我们假设这些变化可能与这些异源蛋白的高产量和可溶性表达有关,我们未来的工作可能集中在Tat标签中碱性氨基酸的数量和分布对异源蛋白表达的影响上。
SOD是细胞中参与活性氧解毒的主要酶之一,在新陈代谢中起关键的抗氧化作用。SOD主要有三个家族,在大肠杆菌中由SodA、SodB和SodC代表。在本研究中,总SOD活性测定显示,Tat标签蛋白的总SOD活性显著高于无Tat标签蛋白。有趣的是,SodA的表达水平高于SodB和SodC。我们假设SodA可能是一种多功能蛋白,尤其是在其抗氧化作用方面。此外,我们分析了Tat标签蛋白和无Tat标签蛋白的抗氧化潜力:细菌增殖活性测定显示,随着诱导时间的增加,百草枯处理后,Tat-SodA、Tat-SodB和Tat-SodC的抗百草枯活性分别比SodA、SodB和SodC有所增加。此外,与野生型相比,百草枯处理后ΔSodA突变菌株的生长受到抑制,而ΔSodB和ΔSodC突变体显示正常生长。因此,SodA可能在抗百草枯活性中起重要作用。
mRNA转录水平的调控需要有效的转录起始、延伸和终止,其中转录因子是外源蛋白高产量表达的主要决定因素。有效的转录起始直接影响mRNA转录水平。必须选择强启动子和调控序列以实现高产量异源mRNA表达,通常使用如含有T7启动子的pET系列载体等构建体。有效的转录延伸和终止也是实现外源蛋白高水平表达的关键因素,因为它们影响mRNA转录的完整性和特异性。通常通过删除负调控元件来增加mRNA转录的完整性,而强转录终止可以防止非特异性mRNA转录。mRNA的稳定性及其二级结构也与翻译产物的数量直接相关。因此,我们假设Tat标签蛋白的mRNA转录水平可能发生显著变化,并且这种变化可能直接影响异源蛋白的表达。正如预期,培养3小时后,mRNA转录水平显著增加,培养6小时后略有增加。此外,Tat标签蛋白的表达高于无Tat标签蛋白,表明Tat标签可以促进异源蛋白在大肠杆菌中的高产量和可溶性表达。
现有文献指出,使用原核表达系统时,膜蛋白和富含二硫键蛋白的表达极其有限。如何将含有许多二硫键的包涵体蛋白高效重折叠成其天然构象,目前仍然是一个具有挑战性的问题,并正在成为生物工程领域的瓶颈之一。由于本研究表明添加Tat核心肽可以促进异源蛋白在大肠杆菌中的高产量和可溶性表达,我们还对含有两个二硫键的膜蛋白LSECTin的表达进行了以下实验。初步实验数据显示,Tat-LSECTin主要以包涵体形式在细胞裂解上清液中表达,并且Tat-LSECTin蛋白的表达产量显著高于单独的LSECTin。这些结果支持了Tat核心肽也能促进含有二硫键的膜蛋白的高产量和可溶性表达的观点,并表明添加Tat标签可以促进蛋白质在大肠杆菌中的高产量和可溶性表达,无论它们是膜蛋白还是富含二硫键的蛋白质,从而为大肠杆菌中的异源蛋白表达提供了一种新工具。
我们的数据表明,HIV-1病毒编码的Tat核心肽可以促进异源蛋白在大肠杆菌中的高产量和可溶性表达。表达的Tat标签蛋白主要位于上清液中,并且与未标记蛋白相比表现出增加的酶活性;过表达Tat标签蛋白的细胞与未过表达的细胞相比,抗百草枯活性显著增加。Tat标签蛋白的mRNA转录水平高于无Tat标签蛋白;Tat标签蛋白的α-螺旋和转角显著增加,其β-折叠和无规卷曲含量显著降低。这些结果表明,添加Tat标签可以通过增加mRNA转录水平和改变所得融合蛋白的二级结构来促进异源蛋白在大肠杆菌中的高产量和可溶性表达,并表明在生物技术和生物工程的异源蛋白表达中具有重要应用。
相关新闻推荐
1、嗜碱盐单胞菌菌株生理生化与生长特性、最优发酵条件——讨论、结论
3、大肠埃希菌的噬菌体DK-13生物学特性及对污染猪肉的杀菌作用(二)