研究简介
衰老的进化理论(如Weismann的生殖质理论)认为多细胞生物中生殖系与体细胞的分离导致资源向繁殖倾斜,体细胞逐渐积累损伤而衰老。近年来研究发现,单细胞生物(如酿酒酵母)也存在类似的年龄不对称现象:母细胞通过保留损伤物质(如蛋白质聚集体)使子细胞“rejuvenated”。出芽酵母的sirtuin范式Sir2p是建立细胞年龄不对称性的关键,这包括母细胞中受损和聚集的蛋白质的保留。研究人员通过建立SIR2的全球遗传相互作用网络,鉴定了极化体、Bni1p的formin Bni1p和肌球蛋白运动蛋白Myo2p,这些都是有丝分裂细胞分裂过程中分离蛋白聚集体机制的重要组成部分。研究发现子细胞可以通过极化酶体和对肌球蛋白依赖的极化聚集体流入母细胞室来清除损伤。Sir2p在细胞骨架功能和极性中的作用与sir2D细胞中CCT伴侣蛋白在折叠肌动蛋白中受损有关。我们讨论这些发现,基于近期模型假设极性可能进化出以避免克隆衰老,通过建立衰老(类体体)和复活(类胚芽)谱系。
Bioscreen全自动生长曲线分析仪的应用
Bioscreen全自动生长曲线分析仪被用于验证遗传互作关系的可靠性和量化突变体的适应性缺陷。微培养实验以三份形式进行,使用Bioscreen C设备将野生型、单突变体(如sir2Δ)对双突变体(如sir2Δbni1Δ)在液体培养基中培养,Bioscreen每20分钟自动检测一次OD值,持续47小时,生成生长曲线。。通过比较不同菌株的生长速率、延滞期和最大生长量,计算“对数菌株系数”(Logarithmic Strain Coefficient,LSC),并利用t检验和阈值检验(LSC差异需超过3倍标准误)判断相互作用的显著性。Bioscreen还用于评估突变体对肌动蛋白解聚药物Latranculin-B的敏感性。在含15μM Latranculin-B的培养基中,sir2Δbni1Δ双突变体生长显著受抑,进一步支持Sir2在肌动蛋白功能中的作用。
实验结果
通过系统性遗传学筛选与功能分析,揭示了Sir2蛋白、极性体(polarisome)和肌球蛋白Myo2p在蛋白质聚集体不对称分离与逆向运输中的核心作用,并提出了一个整合分子机制与进化意义的模型。Sir2缺失导致CCT伴侣蛋白乙酰化水平升高,使其折叠肌动蛋白的效率降低,进而破坏肌动蛋白细胞骨架的完整性。这解释了Sir2与肌动蛋白相关基因(如ACT1、MYO2)的遗传互作,以及sir2Δ细胞对Latranculin-B的敏感性。极性体组分(Bni1p、Bud6p等)通过核化肌动蛋白缆,形成由子细胞指向母细胞的定向流动,确保聚集体在细胞分裂时被保留于母细胞。子细胞内的聚集体可沿肌动蛋白缆逆向流动至母细胞,该过程依赖Bni1p和原肌球蛋白(Tpm1p),但不需要Arp2/3复合物。这表明逆向运输是肌动蛋白缆依赖的定向过程,而非随机扩散。提出了极性最初可能为避免克隆衰老而演化,而非仅用于形态发生。这一模型将Weismann的生殖-体细胞二分法延伸至单细胞生物,说明年龄不对称可能是生命早期演化的关键特征。确立了Sir2-极性体-Myo2p轴在蛋白质聚集体空间质量控制中的核心地位,揭示了细胞通过主动运输机制实现损伤区隔的策略,为理解衰老的进化起源提供了分子基础。
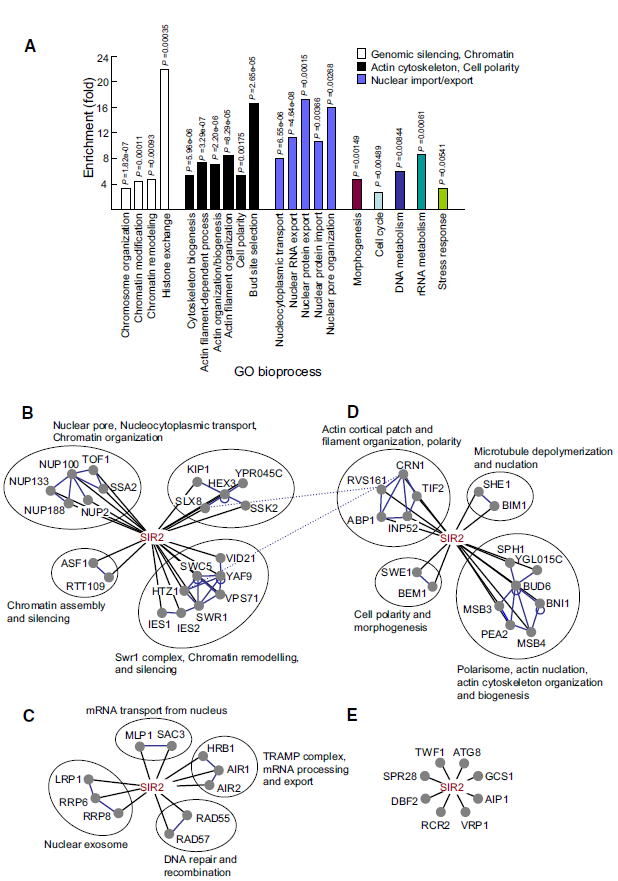
图1、SIR2遗传互作网络的功能富集分析。(A)通过超几何分布分析(p<0.01),确认的SIR2合成致病相互作用在基因本体生物过程组中显著富集,主要涉及染色质沉默、核质运输和肌动蛋白细胞骨架功能。(B-E)基于已知物理相互作用(蓝色边)将SIR2互作基因分组显示。黑色边表示本研究发现的SIR2与复合物间的遗传相互作用。
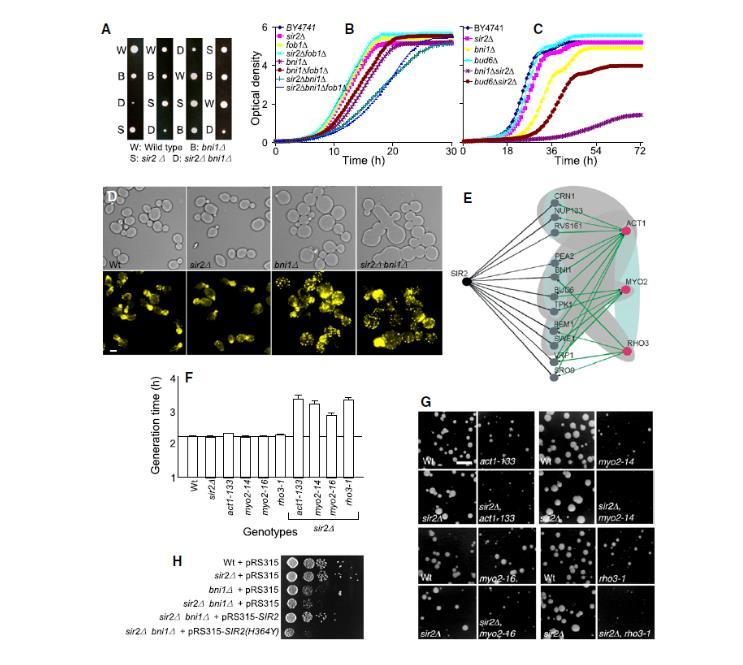
图2、SIR2与极性和肌球蛋白马达machinery的遗传相互作用(A)四分体分析证实sir2Δbni1Δ双突变体的合成致病表型。(B)通过PCR敲除构建的sir2Δbni1Δ突变体在液体培养中显示生长缺陷(深绿线),引入fob1Δ突变(浅蓝线)证明该缺陷与ERC积累无关。(C)sir2Δbud6Δ(棕线)或sir2Δbni1Δ(紫线)对肌动蛋白解聚药物Latranculin-B敏感性增加。(D)sir2Δbni1Δ双突变体出现肌动蛋白斑块聚集和形态异常(上图:明场,下图:罗丹明-鬼笔环肽染色)。比例尺=2μm。(E)遗传互作重叠与物理互作数据预测SIR2与ACT1、MYO2、RHO3的相互作用。黑色边为本研究发现的SIR2互作,绿色边为已知与ACT1、MYO2、RHO3的遗传互作。(F-G)SIR2与温度敏感突变体(act1-133、myo2-14、myo2-16、rho3-1)的合成致病互作。(F)比较30°C或34°C下的世代时间。(G)随机孢子分析结果。比例尺=4mm。(H)野生型Sir2p可抑制sir2Δbni1Δ对Latranculin-B的敏感性,而催化失活突变体(H364Y)无此功能。
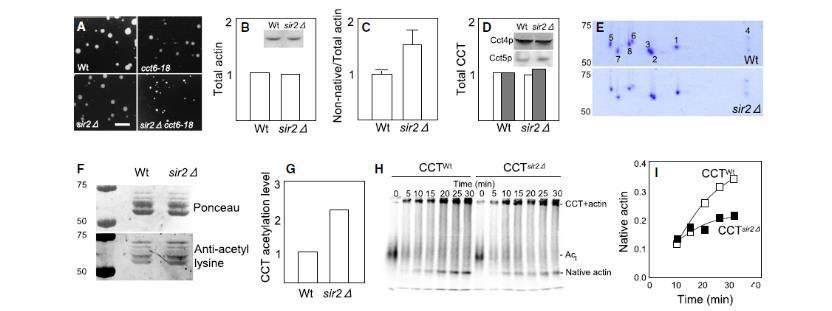
图3、Sir2p缺失细胞天然肌动蛋白减少且CCT活性降低(A)随机孢子分析显示SIR2与CCT(cct6-18)在30°C下的遗传互作。比例尺=4mm。(B)Western blot显示sir2Δ与野生型细胞总肌动蛋白量无差异。(C)DNaseI结合实验表明sir2Δ细胞中非天然肌动蛋白比例增加。(D)CCT亚基(Cct4p、Cct5p)水平在sir2Δ中正常。(E)等电聚焦电泳显示CCT亚基组成无变化。(F)sir2Δ细胞中CCT乙酰化水平升高(上:蛋白负载对照,下:抗乙酰化赖氨酸信号)。(G)抗乙酰化赖氨酸信号量化结果。(H-I)sir2Δ来源的CCT折叠活性降低。放射性标记的β-肌动蛋白与纯化CCT孵育后,通过天然电泳分析折叠效率(I为天然肌动蛋白生成量)。
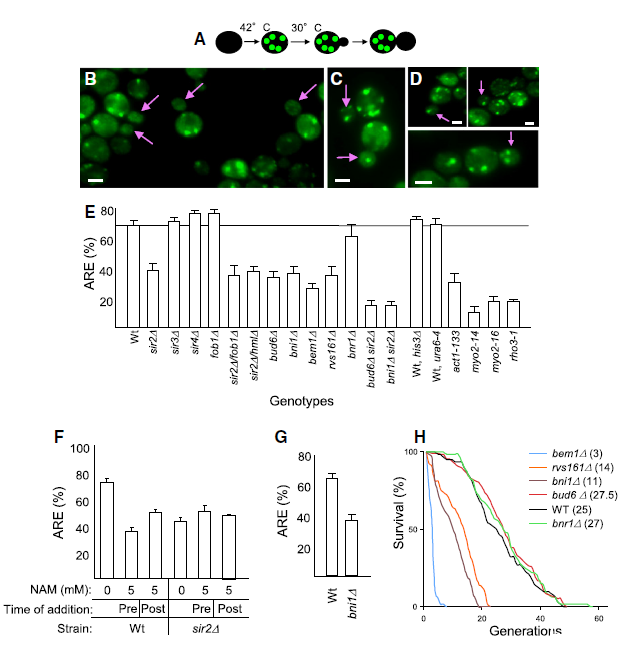
图4、极性和肌球蛋白马达machinery参与蛋白质聚集体的有丝分裂分离。(A)聚集体保留效率实验设计示意图。(B-D)野生型(B)、sir2Δ(C)、bud6Δ(D)细胞胞质分裂期间聚集体分布。比例尺=2μm。(E)不同突变体的聚集体保留效率。(F)Sir2p抑制剂烟酰胺处理降低野生型细胞的保留效率。(G)中年母细胞(平均7代)中衰老诱导的聚集体也依赖Bni1p分离。(H)极性突变体(除bud6Δ外)复制寿命缩短。
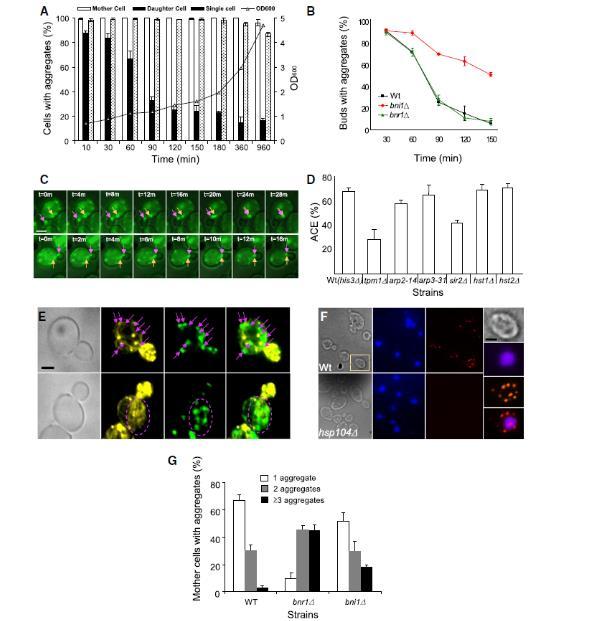
图5、子细胞通过逆向运输在胞质分裂过程中清除蛋白质聚集体。(A)38°C热激16小时后,子细胞聚集体随时间消失,但游离单细胞中重新出现。(B)极性体形成素Bni1p(非Bnr1p)为子细胞清除聚集体所必需。(C)实时成像显示子细胞聚集体向母细胞逆向移动(粉箭头)并与母细胞聚集体(黄箭头)融合。比例尺=2μm。(D)TPM1突变(非ARP2/ARP3)或SIR2缺失会降低逆向清除效率。(E)Hsp104p聚集体定位于肌动蛋白缆富集区(紫箭头)。比例尺=2μm。(F)邻近连接实验检测Hsp104p与肌动蛋白的互作。比例尺=2μm。(G)bnr1Δ突变体减少母细胞内核聚体的融合。
总结
本研究通过系统分析酵母Sir2蛋白的遗传互作网络,揭示了极性体(polarisome)、形成素Bni1p和肌球蛋白Myo2p在细胞有丝分裂过程中介导蛋白质聚集体不对称分离的关键作用,并发现了一种由极性体依赖的逆向运输机制,使子细胞能够清除自身损伤。研究通过整合遗传学、生物化学和活细胞成像,阐明了Sir2-极性体-Myo2p轴在蛋白质聚集体不对称分离中的核心作用,并揭示了子细胞逆向清除损伤的新机制,为理解细胞衰老的进化起源提供了重要依据。研究提出极性最初可能为避免克隆衰老而演化,而非仅用于形态发生。这一模型将Weismann的生殖-体细胞二分法延伸至单细胞生物,说明年龄不对称可能是生命早期演化的关键特征。
Bioscreen全自动生长曲线分析仪被用于验证遗传互作关系的可靠性和量化突变体的适应性缺陷,是支持关键结论的重要实验工具。通过高通量生长分析,揭示Sir2与极性体、肌动蛋白相关基因的协同作用,为阐明蛋白质聚集体不对称分离的分子机制提供关键证据。研究确立了Sir2-极性体轴在空间质量控制中的核心地位,为理解细胞衰老的演化机制奠定了实验基础。
相关新闻推荐
1、重组大肠杆菌E.coli B0013-1050的琥珀酸生成新途径,L-苹果酸产量提高到14 g/L
2、微生物生长曲线分析仪:克氏锥虫在媒介肠道内的定植策略与生存智慧(六)