结果
嗜麦芽寡养单胞菌编码具有多种结构域组成的多个组氨酸激酶。由于嗜麦芽寡养单胞菌ATCC 13637的完整基因组尚未测序,本研究中使用嗜麦芽寡养单胞菌K279a基因组作为参考基因组。根据原核生物双组分系统数据库,该基因组包含60个HK基因,其中16个是混合HK类型,每个都包含一个额外的接收结构域作为磷酸化受体。独立的数据库搜索还显示Smlt2710是一个HWE HK(一组在推测的N和G1框内分别具有特征性组氨酸和色氨酸-谷氨酸残基的HK组)。此外,Smlt4131含有一个具有保守N、G1和G2框的ATP酶结构域,但缺乏典型的HK的H框,表明它是一个退化的HK。Smlt2710和Smlt4131也被纳入分析,因此本研究共检查了嗜麦芽寡养单胞菌的62个推定HK基因(图1)。
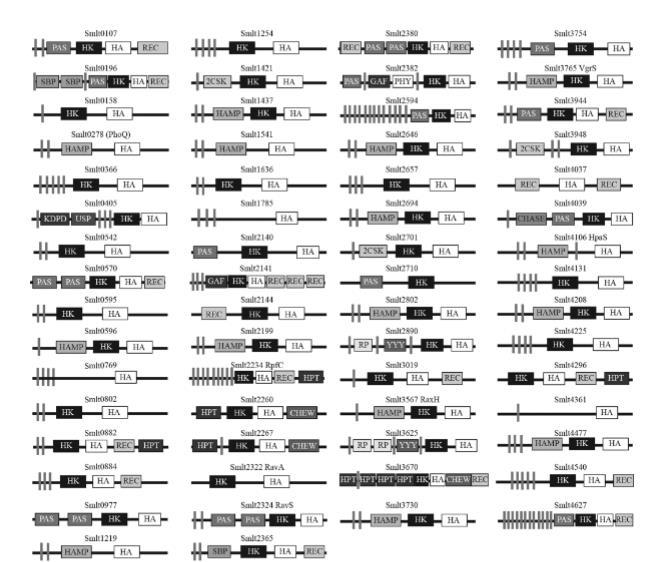
与嗜麦芽寡养单胞菌的近缘物种黄单胞菌属成员的HK基因比较显示,黄单胞菌属中所有与毒力相关的HK的直系同源物,包括RpfC(Smlt2234)、VgrS(或ColS,Smlt3765)、PhoQ(Smlt0278)、RavS(Smlt2324)、RavA(Smlt2322)、RaxH(Smlt3567)和HpaS(Smlt4106),也存在于嗜麦芽寡养单胞菌K279a中(图1)。由于嗜麦芽寡养单胞菌不是植物病原菌,这些与毒力相关的HK在两个细菌类群(寡养单胞菌属和黄单胞菌属)之间的信号检测可能表现出细微的功能分化。这些HK基因中的大多数(56个基因)在基因组附近至少含有一个RR基因。例外的是Smlt0107、Smlt0570、Smlt3019、Smlt3625、Smlt3730和Smlt4540;这六个HK是孤儿HK,它们的同源RR可能缺失或位于其他基因组位点上。
HK基因的系统插入失活和突变体的表型鉴定。为了获得嗜麦芽寡养单胞菌ATCC 13637中HK基因的突变体文库,我们最初旨在构建包含HK基因截短DNA序列(300至450 bp)作为插入片段的重组自杀载体(pK18mob)。在嗜麦芽寡养单胞菌K279a基因组中预测为HK的62个基因中,获得了60个重组载体(见补充材料中的图S1)。两个基因,Smlt1219和Smlt4540,未能通过各种引物进行PCR扩增,表明它们不存在于嗜麦芽寡养单胞菌ATCC 13637基因组中。从这些自杀载体开始,共获得了51个插入突变体,并通过PCR进行了验证(表1)。

尽管付出了巨大努力,但11个基因Smlt1219,Smlt1437,Smlt1541,Smlt2322[ravA],Smlt2382,Smlt2646,Smlt3765[vgrS],Smlt3948,Smlt4131,Smlt4540,和Smlt4627)被证明很难敲除。这些基因在嗜麦芽寡养单胞菌ATCC 13637中可能是必需的,或者它们的失活可能导致在本研究使用的培养条件下严重的生长缺陷。

在丰富NYG平板(1.5%琼脂)和半固体NYG平板(0.1%琼脂)上分别观察了HK基因突变体的菌落形态和游动性(图2),并测量了所有突变体在NYG培养基中的生长曲线(见补充材料中的图S2)。五个基因(Smlt0769,Smlt0882,Smlt0977,Smlt1636,和Smlt1785)的突变体生长速度明显慢于野生型菌株或其他突变体。Smlt0882是肠杆菌荚膜合成中RcsC的直系同源物,其突变通常影响细胞包膜应激反应。Smlt1636在其他细菌中的直系同源物尚未被研究。然而,这两个基因(Smlt0882和Smlt1636)的失活也降低了细菌的游动性(图2C),这可能是由它们的低生长速率引起的。图2C和补充材料中的表S1显示,除了这两个基因突变体外,另外七个突变体也表现出游动能力下降;它们是Smlt1785、Smlt2260(cheA)、Smlt2267(cheA2)、Smlt2324(ravS)、Smlt2380、Smlt2710和Smlt3944突变体。在这些基因产物中,Smlt2260和Smlt2267分别是大肠杆菌生物传感器CheA和CheA2的直系同源物,表明它们参与趋化性调控。Smlt2324(RavS)、Smlt2380、Smlt2710和Smlt3944是含有PAS结构域的HK,负责检测氧气、氧化还原电位或光,从而表明这些信号对嗜麦芽寡养单胞菌的游动性至关重要。
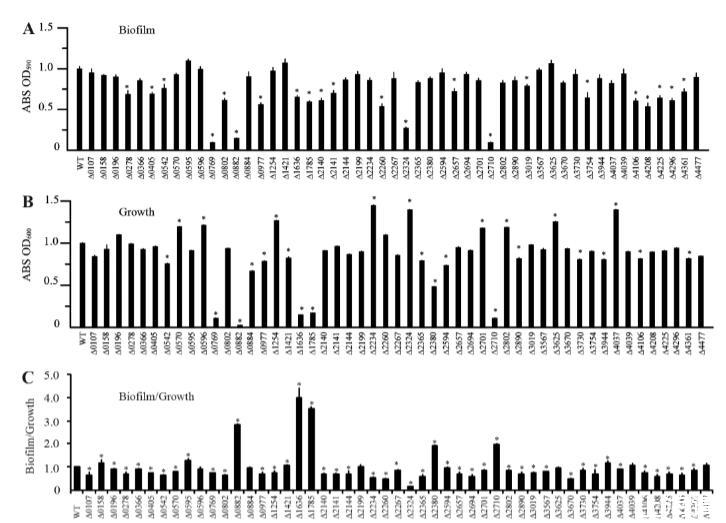
嗜麦芽寡养单胞菌生物膜缺陷突变体的系统筛选。为了研究HK基因突变体的生物膜形成能力,收集在丰富NYG培养基中过夜培养的细菌细胞,进行洗涤,并将各自的浓度调整至OD600为0.4。将等体积的细菌培养物(20微升)接种到96孔板中,每孔含有180微升新鲜NYG培养基。然后将板在28°C孵育5小时,并使用传统的结晶紫染色方法计算生物膜量(图3A)。同时,通过测定每种培养物在OD600的吸光度值来评估孔中的细菌生长(图3B)。为了估计相对于细菌生长的生物膜形成,计算了生物膜量与浮游细菌(OD600)的比率(图3C)。结果显示,一些突变体(ΔSmlt0596,ΔSmlt0884,ΔSmlt3625,和ΔSmlt4477)的生物膜与生长比率与野生型菌株相比没有显著改变。九个突变体(ΔSmlt0158,ΔSmlt0595,ΔSmlt0882,ΔSmlt1421,ΔSmlt1636,ΔSmlt1785,ΔSmlt2380,ΔSmlt2710,和ΔSmlt3944)的生物膜与生长比率显著增加,而其他38个突变体的比率降低。值得注意的是,Smlt2234(rpfC直系同源物)的插入失活导致生物膜与生长比率显著降低(图3C),但无论生长情况如何,其生物膜量并未降低(图3A)。这与黄单胞菌属成员的情况截然不同,后者的rpfC基因突变已被证明会导致细胞间通信和生物膜发育的明显缺陷。
相关新闻推荐
1、RNA聚合酶(RNAP)突变体影响福平(Rif)杀菌/抑菌效应