3.2. 升高的Mg2+离子水平消除TMP介导的生长抑制
上述结果支持PknB_PASTA结构域对PknB活性的功能重要性。PknB_PASTA的结构研究表明,该结构域可以二聚化,因为不同PASTA结构域之间存在构象灵活性。有人提出,假定配体的结合可以诱导激酶结构域的二聚化以及随之而来的PknB的反式激活。因此,PASTA结构域可以作为信号分子的受体并激活细菌复苏和生长,采用先前为枯草芽孢杆菌孢子萌发提出的机制。缺少功能性激酶结构域的PknB_PASTA结构域可能与天然PknB竞争配体,并扰乱PknB介导的信号通路。
因此,我们研究了来自生长分枝杆菌的培养上清液是否会消除由TMP过表达引起的生长缺陷。事实上,通过添加培养上清液以剂量依赖的方式消除了生长抑制。进一步的测试表明,活性化合物是一种耐热的低分子量化学物质,并且在暴露于细菌之前就存在于Sauton培养基中。最终,我们证明活性实体是来自MgSO4或MgCl2的Mg2+。提高的Mg2+浓度可以完全消除PASTA结构域过表达引起的抑制,并且条件培养基的剂量依赖性效应在很大程度上是由于Mg2+浓度增加所致。从较低MgSO4培养基中获得的培养上清液对对照和TMP菌株仅具有中度的生长刺激作用。增加的Mg2+浓度不影响tmp的表达和TMPH的产生。镁也改善了PknB1和PknB5菌株的生长。
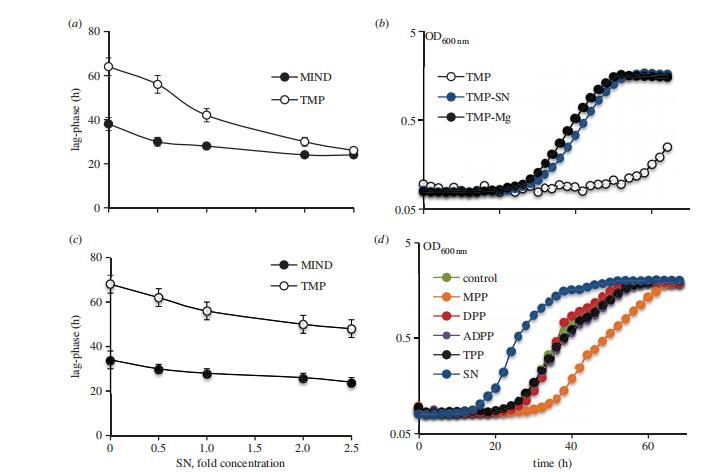
图5. 培养上清液与高浓度 Mg2þ 可消除PknB-pasta过表达引起的抑制效应。(a)培养上清液以剂量依赖性方式消除PknB_TM-pasta过表达对生长的抑制作用。添加培养上清液的相对浓度以‘倍数浓度’表示,1倍值对应未稀释培养上清液。表观滞后期定义为培养物OD600 nm达到0.1所需时间。(b) Mg2þ(10 mM)对 TMP 生长抑制的缓解效果与未稀释培养上清液相当。(c)低浓度 Mg2þ 培养上清液对 TMP 菌株表观滞后期的影响。(d)合成胞壁肽对耻垢分枝杆菌生长的影响。将处于早期稳定期的分枝杆菌用Sauton培养基洗涤两次,取5×10⁶菌体接种于含培养上清液或合成胞壁肽(终浓度10 mM)的Sauton培养基中。MPP:MurNAc五肽;DPP:GlcNAc-MurNAc肽; ADPP :GlcNAc-1,6无水胞壁酰基五肽;TPP:四糖肽;SN:50%(v/v)培养上清液。GlcNAc:N-乙酰葡糖胺;MurNAc:N-乙酰胞壁酸;五肽:L-Ala-g-D-Glu-m-DAP-D-Ala-D-Ala。(b,d)数据为两次独立实验中5次重复的平均值,标准差≤平均值的10%(为清晰起见未显示)。SN:培养基中添加培养上清液。
为了研究PknB_PASTA结构域是否能够结合Mg2+离子,我们使用了NMR方法。用增加量的MgCl2滴定15N标记的重组PknB_PASTA样品。离子与侧链或蛋白质主链的结合会在HSQC实验中测量的1H-15N共振上产生化学位移扰动。然而,在添加MgCl2或EDTA后,在PknB_PASTA中未观察到化学位移扰动,表明蛋白质和Mg2+离子之间缺乏相互作用,并且向重组PASTA中添加MgSO4不会改变重组PASTA的寡聚状态。这些结果表明,Mg2+对TMP介导的生长抑制的改善作用是间接的。
3.3. 胞壁肽不能消除TMP介导的生长抑制
在单独的实验中,我们测试了胞壁肽对TMP菌株生长抑制的影响。我们使用了用各种胞壁质酶消化的大肠杆菌肽聚糖、超声处理或RpfB消化的耻垢分枝杆菌肽聚糖,以及合成胞壁肽。我们仅观察到在相对高浓度的消化或超声处理肽聚糖存在下,TMP菌株的滞后期持续时间从65小时轻微减少到52小时。观察到的滞后期减少不依赖于所用肽聚糖的类型。先前用于枯草芽孢杆菌孢子萌发的合成胞壁肽在任何测试浓度下都不能减少生长抑制,并且不能刺激耻垢分枝杆菌的生长。在其他测试的二价金属中,Ca2+改善了TMP的生长;然而,由于其溶解度差,不能在5 mM以上的浓度使用。由于已知Mg2+离子能稳定细胞壁,并且常用于培养具有缺陷细胞壁的细菌,我们研究了TMP菌株是否比对照对各种抗菌剂更敏感。