SDS-PAGE和Western印迹。 从在BHI培养基中指数生长期中期收获的细胞沉淀制备变形链球菌的蛋白质提取物,并用Tris缓冲盐水洗涤两次。通过在玻璃珠存在下,在SDS煮沸缓冲液中使用珠磨机进行机械破碎,如先前描述制备蛋白质提取物。也将细菌细胞悬浮于4% SDS中,并在室温下孵育1小时以提取表面相关蛋白质。离心后,上清液用作4% SDS提取物。蛋白质通过SDS-聚丙烯酰胺凝胶电泳在含有4.5%堆积胶的10%聚丙烯酰胺凝胶中分离,如Laemmli所述。然后蛋白质用考马斯蓝染色或印迹到Immobilon-P膜上,并进行Western印迹分析的标准技术。膜与针对全长纯化重组AtlA产生的抗-630D1多克隆抗血清一起孵育。通过二辛可宁酸测定确定样品中的蛋白质浓度。感兴趣的条带从染色凝胶中切下,并送往Donald Danforth植物科学中心的蛋白质组学和质谱设施进行鉴定。
转录分析。 通过逆转录酶PCR检查两个基因共转录的潜力。通过实时RT-PCR定量mRNA水平。如先前描述进行RNA提取、RT-PCR和实时RT-PCR,并分析和标准化数据。用于逆转录反应和实时PCR的引物显示在表1中。
结果
atlA操纵子。 我们先前证明变形链球菌atlA基因从至少三个启动子转录,作为四基因操纵子的一部分。在atlA区域的进一步表征中,我们鉴定了一个基因,位于atlA起始密码子上游249 bp,此处显示能够与atlA共转录。这一发现表明SMu0629基因和atlA的基因产物之间可能存在功能联系。
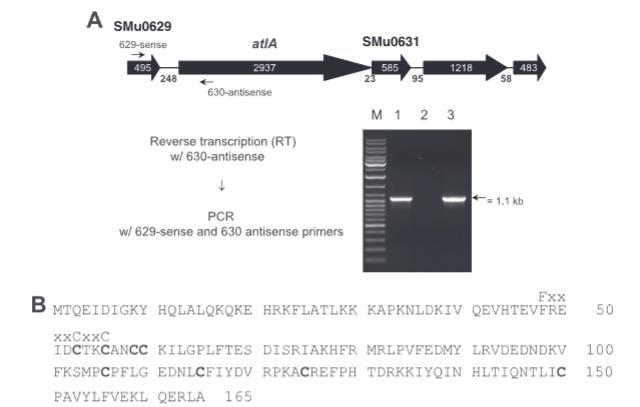
图1. 变形链球菌SMu0629基因座的转录分析。(A)atlA区域示意图及RT-PCR分析。示意图上方的基因命名与编号基于变形链球菌UA159的基因组序列信息。箭头指示转录方向,箭头内部及开放阅读框之间的数字分别表示开放阅读框和基因间区的碱基对长度。使用反向引物(630-antisense)进行逆转录后,采用629-sense和630-antisense引物组进行PCR扩增。引物退火位点如图示。PCR产物在Tris-乙酸-EDTA凝胶中的电泳结果如下:M泳道为分子量标准;1泳道为RT-PCR产物;2泳道为未加逆转录酶的阴性对照;3泳道为UA159染色体DNA的PCR阳性对照。(B)由165个氨基酸组成的SMu0629蛋白序列。该序列含八个可能形成金属结合位点的保守半胱氨酸(加粗显示),以及硫氧还蛋白超家族多个成员活性位点特有的FX4CXXC基序(序列上方标注;第48至56位氨基酸)。
SMu0629基因长495 bp,编码一个保守的假设蛋白,预测分子量为19,504 Da。Pfam搜索显示,残基51至155将SMu0629置于一个未表征的UPF0153蛋白家族中,该家族包含八个保守的半胱氨酸,被提出形成金属结合位点。值得注意的是,SMu0629还包含一个F-X4-C-X-X-C基序。C-X-X-C基序已知是硫醇-二硫键氧化还原酶家族成员的活性位点,这些酶参与多种氧化还原活性,包括蛋白质还原和氧化应激耐受。鉴于AtlA在生物膜形成中的重要作用以及atlA和SMu0629基因产物之间可能存在功能联系,我们探讨了氧气对各种变形链球菌菌株生长和生物膜形成的影响。
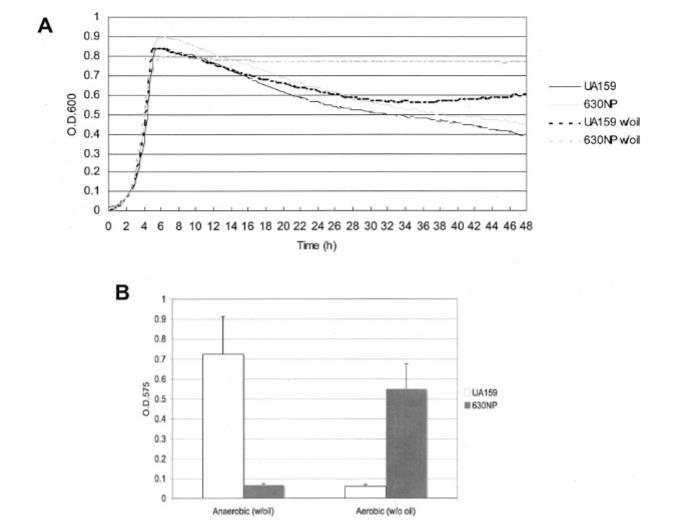
图2. 变异链球菌菌株(野生型与630NP)的生长情况。(A) 在BHI肉汤中的需氧生长通过Bioscreen C系统监测,该系统设置为每30分钟振荡15秒(需氧条件)。对于厌氧生长,在肉汤培养物表面覆盖无菌矿物油(覆油条件)。(B) 变异链球菌UA159和630NP在添加蔗糖的BM培养基中培养48小时形成的生物膜。需氧培养在摇床(150 rpm)中进行,厌氧培养则覆盖矿物油。详见正文。数据代表至少两次独立实验(三次或以上重复)的结果。误差棒表示标准偏差(n=6)。
氧气对自溶和生物膜形成的影响。 为了比较变形链球菌在氧气存在和不存在下的生长,在好氧和厌氧条件下在BHI培养基中监测生长,如材料与方法中详细描述。如图2A所示,野生型和AtlA缺陷菌株的生长速率没有观察到差异。然而,在厌氧条件下,atlA突变体在稳定期晚期显示出对自溶的显著抗性。光学显微镜观察显示,当突变体在好氧条件下生长时,未观察到atlA缺陷菌株特征性的长链形成。
对于体外生物膜测定,微孔板在空气中在旋转摇床上在好氧和厌氧条件下生长。有趣的是,当UA159在通气条件下生长48小时时,在补充蔗糖的BM培养基中形成生物膜的能力显著受损。此外,在相同条件下,atlA突变体比野生型菌株积累了更多的生物膜生物量。当变形链球菌用蔗糖培养时,Gtf酶产生大量水不溶性葡聚糖,促进生物膜的粘附和积累。在补充葡萄糖的BM培养基中的生物膜形成无法研究,因为在这些条件下630NP形成生物膜的能力非常差。氧气对生物膜形成的影响以及通过atlA突变在空气中恢复生物膜形成表明细胞自溶可能受氧气影响,导致变形链球菌形成生物膜的能力变化。我们推断与氧气中生长相关的这些表型可能与SMu0629作为潜在氧化还原酶的功能有关。因此,我们表征了缺乏SMu0629基因的菌株的AtlA生物发生和生长特性。
SMu0629基因突变体的构建和表征。 通过缺失和插入非极性或极性卡那霉素盒破坏SMu0629基因,创建菌株629NP和629P。通过实时RT-PCR确认SMu0629基因中的非极性插入允许高效通读到atlA。从菌株629P的总RNA测量从atlA紧邻上游转录产生的atlA mRNA,而不是作为与SMu0629基因共转录的结果。如图3A所示,629P中的atlA mRNA比629NP或野生型少约50%,证实了基因可以共转录,并为SMu0629和AtlA之间可能的功能联系提供了证据。
在BHI培养基中这些菌株的生长速率没有观察到明显差异。还值得注意的是,在629NP或629P突变体中未观察到atlA缺陷突变体特征性的聚集和链长增加。有趣的是,SMu0629基因突变体在补充葡萄糖的BM培养基中显示出生物膜形成的显著增加,与野生型菌株相比,24小时后增加25%,48小时后增加55%。然而,在BM-蔗糖中未观察到生物膜形成的明显差异。
相关新闻推荐
1、猫疱疹病毒Ⅰ型毒株HRB2019分离培养、增殖与鉴定方法(二)