摘要
诸如恶臭假单胞菌之类的微生物在有机废物和有毒化合物的矿化中扮演重要角色。为了全面准确地阐明恶臭假胞菌中尼古丁降解的关键过程,我们利用基于质谱的谱图计数技术,测量了在以尼古丁或非抑制性碳源甘油生长的恶臭假单胞菌S16中的差异蛋白质丰度水平。计算机分析突显了参与尼古丁降解功能途径的蛋白质的显著聚类。通过定量逆转录PCR分析了差异表达基因的转录调控。我们观察到以下关键结果:(i) 包含1,292个观测蛋白质的蛋白质组提供了尼古丁代谢所涉及酶的详细视图。这些蛋白质可被归类到转运、解毒和氨基酸代谢等功能组。在尼古丁培养基和甘油培养基中生长的细胞,其胞质蛋白质模式存在显著差异。(ii) 3-琥珀酰吡啶转化为6-羟基-3-琥珀酰吡啶的关键步骤是由一个多酶反应催化的,该反应包含一个钼蝶呤结合氧化酶(spmA)、一个钼蝶呤脱氢酶(spmB)和一个(2Fe-2S)结合铁氧还蛋白(spmC),并以钼蝶呤胞嘧啶二核苷酸作为辅因子。(iii) 克隆了一个新型尼古丁氧化还原酶(nicA2)的基因,并对重组蛋白进行了表征。本研究中鉴定的蛋白质和功能途径是降解环境有毒化合物的有吸引力的靶标。
引言
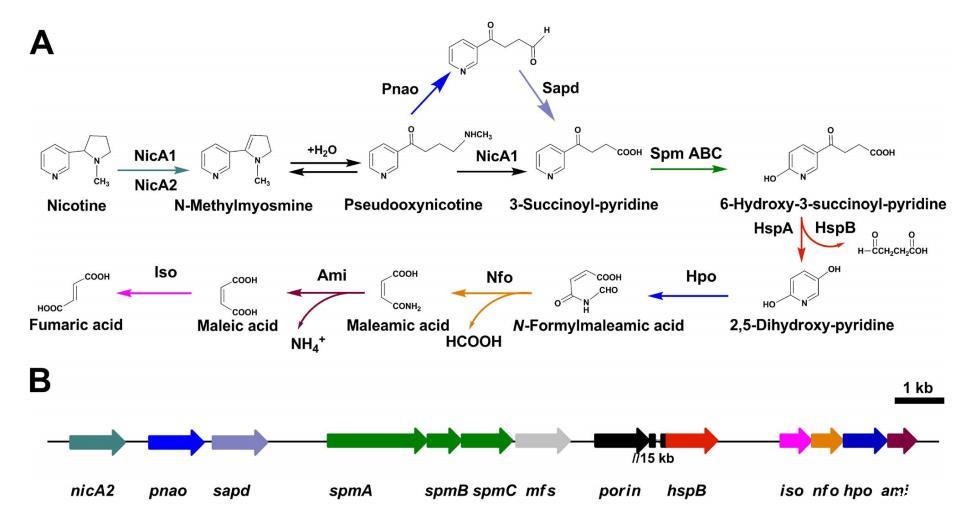
恶臭假单胞菌属于假单胞菌属的非致病性成员,能够代谢多种有机物质图1。HSP羟化酶(HspA或HspB)将HSP转化为2,5-二羟基吡啶(DHP)和琥珀酸半醛[12,13]。恶臭假单胞菌S16通过中间体N-甲酰基马来酰胺酸、马来酰胺酸和马来酸将DHP转化为富马酸,这是尼古丁降解途径的后期步骤(图1)。相关的四个基因,即2,5-DHP双加氧酶基因(hpo)、N-甲酰基马来酰胺酸脱甲酰基酶基因(nfo)、马来酰胺酸酰胺酶基因(ami)和马来酸顺反异构酶基因(iso)已在大肠杆菌中克隆并表达[14,15]。通过计算工具(如Glimmer和Blast同源性搜索)已生成了S16菌株整个基因组的初步注释[16]。因此,有必要鉴定那些通过同源性建模不明显的、能够降解尼古丁的其他蛋白质和途径,并且应该可以使用蛋白质组学方法来搜索涉及尼古丁分解代谢的调控和分子机制的蛋白质。
尽管对尼古丁降解的科学重要性的认识日益提高,但关于编码SP羟基化和细菌中尼古丁转化的关键基因知之甚少。在过去几年中,人们通过基因组文库筛选和野生型酶纯化付出了巨大努力来鉴定SP羟基化的关键基因。不幸的是,这些努力未能鉴定出任何与SP羟基化相关的基因。进展可能需要开发综合的实验方法。通过蛋白质组学方法,可以观察生物体合成和利用的整套蛋白质产物[17]。它可以帮助我们准确确定ORF的边界和枚举,并验证那些不能基于同源性很好确定的未知ORF。
在本研究中,我们分析了S16菌株的胞质蛋白质模式,该菌株含有尼古丁降解的必需基因,使用多维液相色谱-串联质谱方法和分子遗传学。该蛋白质组信息用于补充相应蛋白质子集的基因诱导和表达数据。通过MS/MS谱分析,以最佳得分(命中数>=5, 唯一性>=2)鉴定出总共1,292个蛋白质。涉及转运、能量、解毒和应激反应的蛋白质在尼古丁存在下上调。此外,暴露于尼古丁导致膜蛋白上调,包括孔蛋白、外膜蛋白和与溶剂外排泵相关的蛋白质。本研究代表了对尼古丁降解中细胞反应的全局分子表征的重要途径。我们提供了SP羟化酶参与该途径的证据,该酶由一个4.45 kb的基因簇编码。该酶需要钼蝶呤胞嘧啶二核苷酸作为辅因子。此外,我们描述了另一种尼古丁氧化还原酶(NicA2),其与NicA1(先前称为NicA)的氨基酸同一性较低(10.9%)[11]。删除nicA2而非nicA1阻止了恶臭假单胞菌S16对尼古丁的分解代谢,证明了新发现的NicA2的重要性。构建了四个缺失突变体,并表明这些酶NicA2、Mfs、Pnao和SpmABC对于尼古丁降解是必需的。据我们所知,这是首次报道在假单胞菌菌株中定量研究尼古丁诱导的全局蛋白质和mRNA表达变化。我们将比较蛋白质组学数据与分子遗传学相结合的方法揭示了可能参与尼古丁降解的有趣的假单胞菌特异性蛋白质。
尼古丁和甘油培养基中恶臭假单胞菌S16尼古丁降解的分子反应的蛋白质组学鉴定

为了揭示尼古丁降解中的分子途径,我们获得了参考条件(甘油作为唯一碳源和(NH4)2SO4作为氮源)与处理条件(尼古丁作为唯一碳源和氮源)之间蛋白质表达变化的全局概览。在LC-MS/MS测量阶段获得了尼古丁和甘油条件之间的直接比较。使用LTQ Orbitrap XL混合FTMS分析对肽段和相应的蛋白质进行鉴定和定量分析。每个条件(尼古丁或甘油作为培养基)测量三个独立运行,共使用六个生物样品。通过MS/MS谱分析鉴定出总共1,292个通过得分(命中数>=5, 唯一性>=2)的蛋白质(表S1)。这1,292个观测到的蛋白质代表了理论蛋白质组(S16菌株的5,218个推定编码序列,[16])的25%。使用全局归一化消除实验误差,并对两组蛋白质之间进行t检验。只有变化倍数>=3倍且p值<0.05的蛋白质才被报告为差异表达蛋白质。根据这些预定义标准,126个推定蛋白质在尼古丁和甘油培养基之间显示出显著的蛋白质丰度变化(表S2和表S3)(图S1和图2)。根据S16菌株的整个基因组和蛋白质组数据[16],提供了恶臭假单胞菌S16环状染色体的图谱,说明了已知基因、预测编码区、基因组岛和差异表达蛋白质的位置。
相关新闻推荐
1、高产磷酸吡哆醛功能性乳酸菌筛选、最适生长温度及生长曲线图绘制(三)