结果
分子鉴定
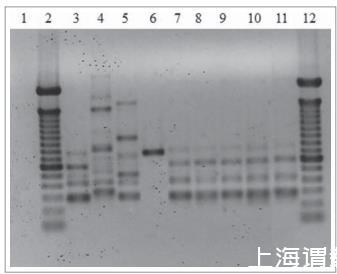
5株分离株的T3B PCR指纹图谱显示300至800 bp的DNA片段,通过与巴西孢子丝菌模式菌株获得的图谱相似性可以进行清晰鉴定(图2)。此外,来自所有患者分离株的图谱呈现100%相似性。
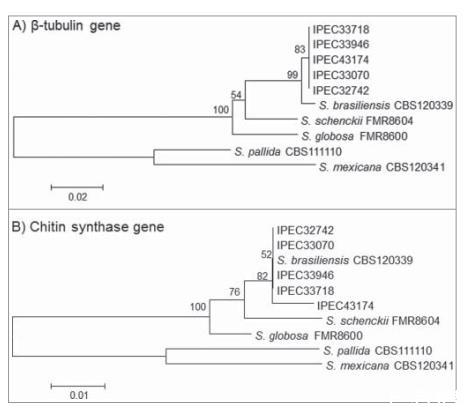
基本局部比对搜索工具(BLAST)分析将分离株的部分β-微管蛋白(β-tub)和几丁质合酶(CHS)序列与NCBI GenBank数据库(AM116954,AM116950,AM116956,AM116946,AM114898,AM117414,AM114889,和AM117417)中的分离株进行比较,以99-100%的相似性将这些分离株鉴定为巴西孢子丝菌。此外,将从所有分离株获得的部分β-tub和CHS序列与GenBank数据库中可用的孢子丝菌复合体参考菌株进行比较和系统发育分析,结果显示与巴西孢子丝菌相似,并且两个基因都显示出高bootstrap支持值(图3A和B)。分离株5的部分CHS序列与前4株分离株相比有一个2-bp的多态性(位置109 A/C和151 G/C),表明该分离株在系统发育分析中存在变异性。获得的β-tub和CHS编码基因序列已存入GenBank数据库,登录号为KM222420至KM222429。
体外实验
为了表征其毒力,进行了使用J774巨噬细胞的体外实验,包括吞噬作用、真菌杀灭和巨噬细胞活力测定。有趣的是,分离株之间没有发现差异(p>0.05),并且使用巴西孢子丝菌模式菌株获得了类似的结果。酵母与巨噬细胞相互作用2小时后的吞噬指数范围从分离株5的89%到分离株4的95%,并且巨噬细胞无法抑制巴西孢子丝菌的生长,18小时后真菌存活率在96%至123%之间变化。在此时间点后,所有条件下的巨噬细胞活力约为90%(范围从分离株5的89%到分离株3的92%)。
大蜡螟感染模型
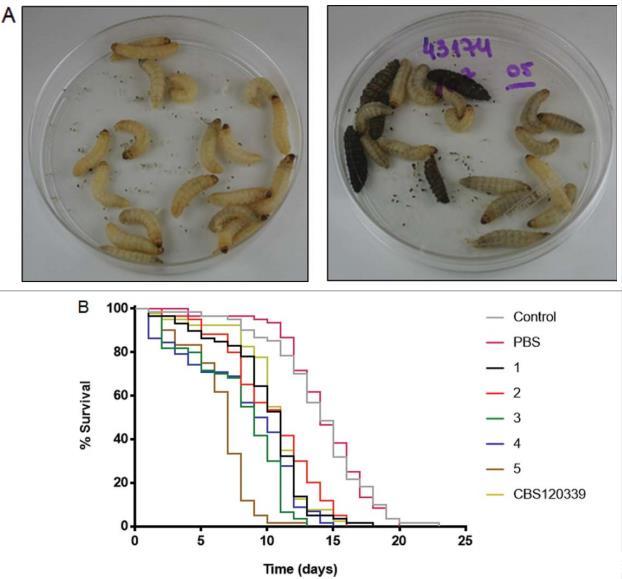
通过用107个酵母细胞攻击幼虫,进行了大蜡螟幼虫的体内实验。与对照组相比,接种酵母的幼虫色素沉着增强(图4A)。获得了3个独立实验的生存曲线。与对照组相比,所有分离株都降低了幼虫的存活率(P<0.0001),但与注射分离株5的幼虫组相比,这种降低始终显著更高。图4B显示了3个实验合并数据的生存曲线,其中分离株5的中位生存期为7天,而其他分离株为9-11天(P<0.0001)。未感染对照组的中位生存期为14天。我们还用104和106个酵母进行了实验,但在这些接种量下,幼虫的存活率与未感染对照组相似(数据未显示)。
在大蜡螟感染后第3天和第6天,真菌负荷没有显著差异。感染后第6天获得的CFU范围从分离株4的1.4 x 107到分离株1的8.2 x 107。
推定毒力因子
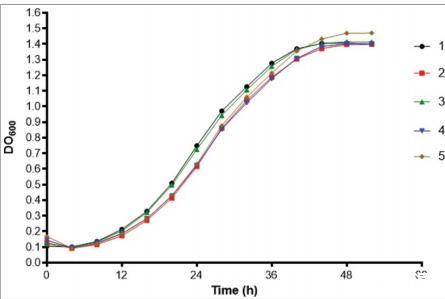
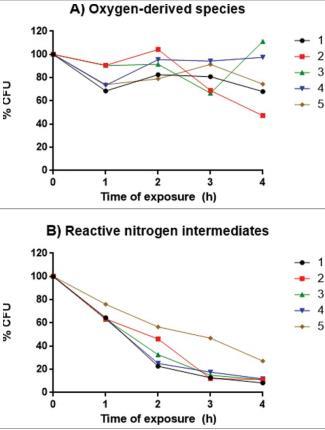
由于我们发现分离株5在大蜡螟中的毒力与其他分离株不同,我们评估了与毒力相关的因素是否可以解释这种变化的原因。在37°C下使用Christensen尿素肉汤测定脲酶产量,所有5株分离株在4天和7天后均为阳性,559 nm处的平均吸光度没有差异。通过测量琼脂上azoalbumin降解晕圈确定的蛋白酶活性范围从0.6到0.76,分离株1的活性最高(p/z=0.6)。该活性与分离株2(p=0.016)和分离株4(p=0.0005)有显著差异。分离株产生的DHN-黑色素比例(通过干重DHN-黑色素/干重酵母比率获得)没有统计学差异。所有分离株在35和37°C下生长,在39°C下均不生长。此外,分离株在BHI培养基中的生长曲线相似,它们之间没有统计学差异(P>0.05;图5)。对于氧源性物质的暴露,分离株2在暴露4小时时生长抑制最高,这与分离株3和分离株4有显著差异(P<0.05)(图6A)。
其他时间间隔的差异不显著。暴露于活性氮中间体降低了分离株的存活率,但分离株5的存活率在检查时间内始终高于其他分离株。然而,这些差异不显著(图6B)。
药敏试验
对所有分离株的抗真菌药物敏感性相似,特比萘芬的MIC为0.06μg/mL,伊曲康唑为1.0μg/mL,酮康唑和两性霉素为2.0μg/mL,伏立康唑≥8.0μg/mL。
讨论
我们的研究重点是对该患者连续分离株的表征,考虑到他在11年期间孢子丝菌病的临床侵袭性和治疗困难,疾病播散到皮肤和骨骼。我们必须指出,缺乏先前治疗在巴西某些人群中非常普遍,原因是农村地区距离诊所遥远、即使像碘化钾这样便宜的药物也负担不起,以及不愿接触医疗系统。尽管患有糖尿病和高血压,该患者之前是健康的,这体现在他作为砖瓦工在physically stressful条件下积极工作的能力,并且为寻找可能的潜在免疫抑制状况进行的测试没有显示异常。初始T细胞的减少可能与慢性感染引起的淋巴细胞持续刺激,诱导其分化有关。
考虑到该患者疾病的慢性和复发性,我们考虑了可能性:1)他可能被不同物种或菌株再感染,或者2)随着时间的推移,感染菌株的毒力发生了变化。然而,我们确认每个分离株都是巴西孢子丝菌。虽然不确定,但发现分离株在指纹PCR中的条带模式没有差异(图2)表明它们是相同的;然而,这并不能排除随着时间的推移被同一菌株再感染,而不是慢性疾病的复发。
分离株5的CHS编码基因的多态性表明几丁质合酶基因发生了改变,这可能通过影响几丁质合成和细胞壁结构来增加毒力。几丁质是真菌细胞壁和隔膜的重要组成部分,存在于所有已知的致病性真菌中,但不存在于人类中。毒力决定因素包括宿主识别和入侵所需因子以及保护免受宿主防御系统侵害的因子。由于细胞壁参与这些过程,它保护真菌细胞免受外部损伤,调节宿主细胞对细胞壁成分的粘附,因此介导免疫反应。因此,细胞壁结构及其组成的任何变化都可能影响真菌毒力。迄今为止,几丁质已与其他真菌的毒力相关,如巴西副球孢子菌,荚膜组织胞浆菌和皮炎芽生菌。几丁质沉积可能影响黑化作用,这是另一种与细胞壁和致病性真菌(包括孢子丝菌)毒力相关的重要因素。因此,几丁质的改变可能导致毒力的改变。
我们的体外巨噬细胞实验没有显示患者分离株之间的毒力差异:巨噬细胞有效地吞噬了酵母,但一旦摄入后无法杀死它们。然而,我们使用大蜡螟幼虫的体内实验导致接种分离株5的幼虫死亡率consistently higher,该分离株是在INI随访5年后(疾病总共11年后)获得的,这强烈表明该分离株的毒力发生了变化。应该指出,大蜡螟模型已成功用于其他真菌和细菌,38-40本研究代表了其在巴西孢子丝菌中的首次使用。大蜡螟模型相对于啮齿动物系统具有优势,例如成本低、易于储存和处理以及更有利的伦理方面。
测定脲酶和DHN-黑色素以及我们评估耐热性的实验显示分离株之间没有差异。蛋白酶活性不均匀,但分离株1测得的较高活性与所有其他分离株相比并不一致,并且似乎并未意味着该分离株毒力增加,如毒力实验所证明。所有分离株的抗真菌药物敏感性相似。将我们患者菌株中发现的MIC与先前发表的里约热内卢孢子丝菌属41和巴西孢子丝菌42的药敏谱进行比较,特比萘芬对分离株产生了最好的体外疗效,而唑类药物和两性霉素B分别具有较差和中等的MIC结果。值得注意的是
7个各分离株酵母或巴西孢子丝菌参考CBS菌株(置于10μL无菌PBS中)的幼虫。幼虫在37°C下维持。N=每组60只幼虫。
尽管特比萘芬的MIC较低,但这是该患者多年来治疗仅取得中等成功的药物。
最后,评估对氧化应激敏感性的测试显示,分离株对活性氮中间体的敏感性高于对氧源性物质的敏感性。值得注意的是,分离株5对活性氮中间体的抵抗力最强。抵抗类似于巨噬细胞溶酶体内发现的不良条件的能力可以解释该分离株毒力更高的原因;然而,我们的结果不能完全得出结论。尽管如此,这一发现可能部分促进了该分离株的更高毒力。i
由于患者自发病以来已退休,远离了他可能感染的环境,我们的数据表明,合理地得出结论他一直是慢性感染。此外,将该患者的分离株鉴定为巴西孢子丝菌有些令人惊讶,因为我们没有发现与接触猫的任何流行病学联系,并且患者居住在里约热内卢孢子丝菌病“带”以外的市镇。
我们的数据支持结论,即该患者在11年期间感染了相同的真菌。尽管其体外对特比萘芬敏感,但巴西孢子丝菌菌株通过逃避他的免疫系统反应有效地留在了患者体内。此外,大蜡螟研究表明,在该患者的疾病过程中毒力发生了演变。我们怀疑寄生虫-宿主关系中存在其他未研究的方面,导致了这种毒力的增加,需要进一步研究来阐明这些因素。我们认为这项体内和体外研究是独特的,因为它证明了巴西孢子丝菌在患有慢性和播散性孢子丝菌病的宿主体内毒力的演变。
致谢
感谢David Alejandro Sanchez在毒力实验中的帮助,感谢Otávio Espíndola博士进行流式细胞术分析,感谢Nelson Lima博士提供墨西哥孢子丝菌菌株。