3.4. 过表达PknB-PASTA增加耻垢分枝杆菌对β-内酰胺类抗生素的敏感性并改变细胞壁蛋白的丰度
我们测试了PknB1、TMP和MIND菌株对各种抗菌剂的最低抑菌浓度。在所有测试菌株中,利福平、乙胺丁醇和链霉素的MIC相同。然而,TMP菌株对几种β-内酰胺类抗生素显著更敏感:美罗培南、氨苄西林和克拉维酸。在亚抑制浓度的克拉维酸存在下,TMP菌株对美罗培南的MIC降至0.15μM以下,而对照菌株为6.5μM。有趣的是,过表达PknB并未改变抗菌敏感性,表明PknB-PASTA结构域本身可能干扰肽聚糖合成,导致对β-内酰胺类更高的敏感性。肽聚糖合成的干扰也可以解释在TMP菌株中观察到的延长的滞后期。为了解决这种可能性,研究了滞后期从MIND和TMP菌株分离的膜蛋白。
表1. PknB-PASTA过表达对耻垢分枝杆菌抗生素敏感性的影响
| 菌株 | 利福平 | 链霉素 | 乙胺丁醇 | 美罗培南 | 克拉维酸 | 美罗培南 + 克拉维酸a | 氨苄西林 |
|---|---|---|---|---|---|---|---|
| MIND | 1.2 | 0.4 | 2.5 | 13 - 19.5 | >1000 | 6.5 - 13.0 | 140 - 170 |
| PknB1 | 1.2 | 0.4 | 2.5 | 13 - 19.5 | >1000 | 6.5 - 13.0 | 140 - 170 |
| TMP | 1.2 | 0.4 | 2.5 | 6.5 | 150 - 200 | 0.13 | 30 - 45 |
| a 克拉维酸的添加浓度为 100 μM。 | |||||||
使用LTQ-Orbitrap-Velos质谱法,我们在两种菌株的膜组分中总共鉴定出1071种蛋白质。TMPH菌株有90种蛋白质是该样品特有的,而MIND菌株仅检测到95种蛋白质。大多数蛋白质在两种菌株中都有鉴定。重要的是,通过检测TMPH菌株中22个独特的片段,证实了膜组分中PknB_PASTA结构域的产生。大多数检测到的蛋白质被归类为功能未知的假设蛋白,而267种被预测为膜或细胞表面蛋白质。这一结果证实提取程序选择性富集了膜蛋白。
在两种菌株的膜组分中都发现了大量涉及金属、营养物质和酶转运的膜蛋白。接下来,使用TMTsixplex试剂盒标记肽段,以比较TMP和MIND中膜蛋白的相对丰度。我们专注于细胞分裂蛋白和参与细胞壁生物合成的酶的分析。大多数细胞分裂蛋白在TMP和MIND菌株中同样存在。然而,CwsA在TMP菌株中更丰富。
表2. TMP与MIND菌株相比细胞壁酶和蛋白质的差异表达| 结核分枝杆菌蛋白 | 耻垢分枝杆菌蛋白 | 蛋白质描述 | 比率 TMP/MIND (log2倍数变化) |
|---|---|---|---|
| Rv0014c | MSMEG_0028 | 带组氨酸标签的TM-PASTA | 5.9 |
| Rv0008c | MSMEG_0023 | 细胞壁蛋白 CwsA | 1.0 |
| Rv3804c | MSMEG_6398 | 分枝酸转移酶 FbpA | 1.0 |
| Rv3790 | MSMEG_6382 | DprE1 十异戊二烯磷酰-β-d-核糖 2'氧化酶 | 0.9 |
| Rv3577 | MSMEG_6071 | 金属-β-内酰胺酶 | 0.9 |
| Rv0129c | MSMEG_3580 | 分枝酸转移酶 FbpB | 0.6 |
| Rv3782 | MSMEG_6367 | 半乳呋喃糖基转移酶 | 0.8 |
| Rv3808c | MSMEG_6403 | 半乳呋喃糖基转移酶 | 0.8 |
| Rv3265c | MSMEG_1826 | dTDP-Rha-a-D-GlcNAc-二磷酸多萜醇, a-3-L-鼠李糖基转移酶 | 0.7 |
| Rv0906 | MSMEG_5638 | 金属-β-内酰胺酶 | 0.6 |
| Rv2518c | MSMEG_4745 | LdtB L,d-转肽酶 | 0.6 |
| Rv0237 | MSMEG_0361 | 糖基水解酶 | 0.6 |
| Rv3330 | MSMEG_1661 | DacB1 D-丙氨酰-D-丙氨酸羧肽酶 | 0.5 |
| Rv2748c | MSMEG_2690 | DNA易位酶 FtsK | 0.5 |
| Rv2171 | MSMEG_4239 | 保守脂蛋白 | -0.8 |
| Rv2721c | MSMEG_2739 | 跨膜富含丙氨酸和甘氨酸蛋白 | -0.5 |
CwsA的确切功能尚不清楚,但已显示它与CrgA相互作用。CrgA仅在TMP的膜组分中检测到,而在MIND中未检测到,因此无法计算其相对丰度。另一个CrgA相互作用伙伴Wag31在两种菌株中以相似水平检测到。有趣的是,TMP菌株有更多参与分枝酸-阿拉伯半乳聚糖生物合成的酶,包括几种分枝酸转移酶和半乳呋喃糖基转移酶。关于肽聚糖相关酶,两种金属β-内酰胺酶、L,D-转肽酶LdtB和D-丙氨酰-D-丙氨酸羧肽酶DacB1的水平在TMP膜组分中增加。完整数据可通过ProteomeXchange获取。总的来说,这些结果可能表明PASTA结构域的过表达影响分枝杆菌细胞壁的生物合成和结构。
3.5. 添加重组PASTA蛋白可消除结核分枝杆菌的生长
上述结果表明,过表达PknB_PASTA结构域可用于抑制医学上重要的病原体结核分枝杆菌的生长并增加对抗菌剂的敏感性。因此,我们研究了PknB_PASTA结构域是否在结核分枝杆菌中具有类似效应。我们将pMind pknB7和空pMind质粒转化到结核分枝杆菌H37Rv中,并研究了所得菌株在四环素存在下的生长。
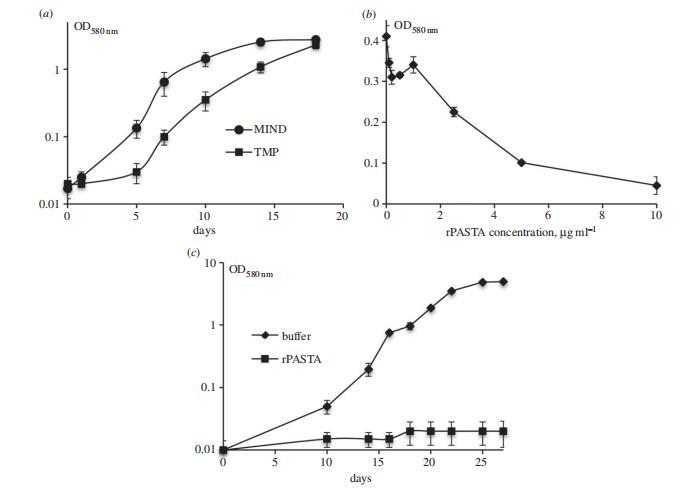
图6. PknB_PASTA抑制结核分枝杆菌生长。(a) PknB_PASTA结构域过表达延迟生长启动。将分枝杆菌(约1×10⁶个细胞/毫升)接种于含潮霉素和四环素的补充7H9培养基中。(b) 不同浓度rPASTA对结核分枝杆菌生长的影响。将分枝杆菌(10³个细胞/毫升)接种于含不同浓度无菌rPASTA的补充7H9培养基中,培养21天后测定光密度值。(c) 添加rPASTA延缓储存结核分枝杆菌的再生长。将结核分枝杆菌(2×10⁴个细胞/毫升)接种于补充7H9培养基中,以缓冲液作为对照组,rPASTA终浓度为10 mg/ml。(b,c) 数据点为3次生物学重复的平均值,误差线表示标准偏差。实验重复三次,图中展示典型实验结果。
如图所示,过表达外部膜锚定的PASTA确实抑制了结核分枝杆菌生长的启动。我们接下来研究了添加仅包含PASTA单元的重组PASTA蛋白是否可能改变生长。我们注意到,向结核分枝杆菌中添加rPASTA在浓度为10μg ml-1时导致生长缺陷,并使表观滞后期从5天增加到14天。这种生长抑制是暂时的,培养物最终产生正常生长,而向对数期培养物中添加重组PASTA对生长没有影响。然而,当分枝杆菌在4°C储存两个月时,rPASTA的生长抑制效应更为明显。
我们发现这些细胞保持了可培养性,并在液体和固体培养基中均产生正常生长。然而,添加浓度为10μg ml-1的rPASTA显著延迟了它们的生长启动。分枝杆菌未被rPASTA杀死,在两个月的培养后产生了正常的稳定期培养物。在所有实验中,对照缓冲液没有任何抑制作用。这些结果表明,PASTA结构域可能直接干扰分枝杆菌的再生长,因此为设计改变细菌再生长和复苏的特异性药物提供了一个可能的靶点。