3.3. φR1-37转录的时间调控
可以通过总结每个样本中比对到噬菌体基因组任一条链上每250 bp的读数,通过总计数(TC)将每个样本中比对到噬菌体的读数彼此归一化,并分别绘制每条链,来可视化感染过程中每个测量点的噬菌体转录(图3A)。这个图谱显示,在早期、中期和晚期阶段,基因转录模式总体稳定。大多数分析的基因在所有研究的时间点都呈现出相当相似的表达模式。只有少数选定的基因在比较感染早期和晚期时显示出表达的明显差异。对单个基因的早期(2-5分钟)、中期(10-21分钟)和晚期(28-49分钟)阶段的平均读数计数的比较表明了四种不同的基因表达模式(图3B)。
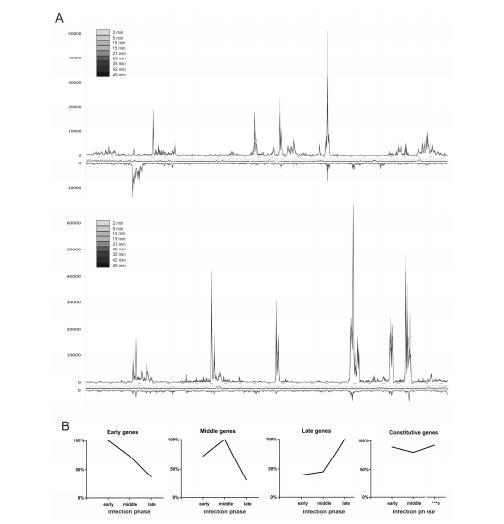
图3. φR1 -37基因组转录在感染周期中的动态变化。图中展示了各时间点时,噬菌体基因组两条链上每250 bp片段的读段数分布(A)。曲线颜色由浅至深的渐变强度对应连续时间点(图A高分辨率图像参见图S4)。 φR1 -37基因表达的不同时序特征(B)。针对每个基因及感染阶段,将最高总基因读段数(TGR)值设定为100%,其余数值按比例调整。
第一组包括92个φR1-37基因,其转录本在感染的前10分钟内积累,因此可归类为早期基因。第二组(8个基因)的转录本丰度在感染后15至28分钟之间达到最大水平,允许归类为中期基因。归类为晚期的第三组94个基因的转录本数量随时间增加,在35至49分钟之间达到峰值。最后,最大的第四组包括147个φR1-37基因,它们在感染后不同时间点的丰度没有显著变化,被归类为组成型表达基因。在367个基因中,有26个由于在这些采样条件下表达水平非常低而被排除。没有观察到基因的基因组位置与表达模式之间存在明确的关联。这些发现与典型的时序模式(即转录本从早期基因向晚期基因转移,并且同一类基因在基因组中成簇排列)完全不同,正如使用RNA-seq或更早的方法在其他几种噬菌体中所观察到的那样。由于大多数φR1-37基因缺乏明确的功能注释,目前甚至无法推测许多组成型表达基因的功能。
在92个早期基因中,87个与已知基因没有相似性。其他早期基因编码一种内切型膜结合裂解性胞壁质转糖基酶(g331)、一种含有Pro-Ala-Ala-Arg(PAAR)重复序列的蛋白质(g142)以及,出乎意料地,两种病毒粒子结构蛋白(g088,g094)。在18个中期基因中,只有一个具有预测功能,即一个推定的RNA结合蛋白(g284)。大多数涉及DNA复制和修复的基因(30个中的22个)被归类为晚期基因。晚期基因(n=94)包含20个编码病毒粒子结构蛋白的基因、4个核苷酸代谢基因和3个RNAP及RNA相互作用基因。有趣的是,大多数编码病毒粒子结构蛋白的基因(70个中的48个)被归类为组成型表达,在整个感染过程中显示稳定的表达。类似地,七个编码DNA复制和修复因子的基因、两个编码ATP酶的基因以及三个编码参与核苷酸代谢的因子的基因被归类为组成型表达(表S2)。此外,两个注释为编码裂解相关蛋白的基因(g084和g289)也属于同一类别,尽管与具有最高TGR值之一的编码裂解蛋白的g331相比,它们的表达水平非常低(表S2)。需要进一步的研究来通过实验确定g084和g289产物的功能以及它们的组成型表达对宿主裂解的可能影响。虽然我们观察到这两个预测参与裂解的基因与标准感染模型相悖,但它可能表明存在类似噬菌体T4中发现的复杂裂解抑制机制。
另一方面,在具有最高TGR值的基因中,我们鉴定了编码病毒粒子结构蛋白(g326,g176,g083,g174,g099)、ATP酶(g304,g061)和裂解蛋白(g331)的基因。没有读数比对到基因g067,g106,g109,g116,g165,g166,g172,g180,g181,g188,g200,g201,g217,g232,g288,g312,g315,g316和g357。关于噬菌体基因表达的详细信息见表S2。
RNA测序数据支持我们先前的Northern印迹结果,提供了φR1-37转录的整体概览。与Northern印迹结果一致,我们在此表明基因g072-g072、g103、g170和g329是晚期表达的,而g231基因表达是组成型的。基因g048-g049、g145、g281和g298的结果存在差异,可以用它们的转录本TGR值非常低来解释。这些转录本丰度低,以及噬菌体转录本在早期时间点普遍丰度较低,使得Northern印迹分析具有挑战性。在这种情况下,高灵敏度的RNA测序被证明是更可靠的方法。
早期研究表明,编码与细菌RNAP β和β'亚基相似的蛋白质的基因在所有已知的φKZ相关噬菌体中都是保守的。在φKZ中,四个编码组成型病毒RNAP的基因是中/晚期基因,而编码非病毒粒子RNAP的基因在早期表达。在体外,φKZ的RNAP复合物以抗利福平的方式从晚期噬菌体启动子起始转录。此外,φKZ RNAP不具备细菌、古菌或真核生物聚合酶特征性的组装和启动子特异性亚基。我们的研究表明,在φR1-37中,编码RNAP亚基的g274、g102和g261基因是晚期且高表达的。其余三个(g178、g231和g099)从感染的最早点开始组成型表达,并且其特征是转录本丰度中等至高。先前假设φR1-37在感染后早期将β和β'亚基引入宿主细胞以接管转录。有可能,类似于φKZ,φR1-37噬菌体强烈依赖于其自身的RNAP,并在整个感染过程中使用它。
相关新闻推荐
3、生长曲线分析仪解析希瓦氏菌呼吸性硝酸盐铵化途径的遗传与生理机制(一)