HdiR自切割分析
在pH 6.0-10范围内使用含有Bis-Tris、Tris-HCl或甘氨酸的缓冲系统研究His6-HdiR的自切割。简言之,每个含有纯化His6-HdiR的反应在室温孵育16小时。反应产物在4-12% SDS-PAGE中分离,随后用考马斯亮蓝R-250染色,或转移到硝酸纤维素膜上,用His6-HdiR抗体和HRP偶联的山羊抗兔IgG进行Western印迹。使用SuperSignal West Dura Extended Duration kit进行检测。在GS-525分子成像系统上扫描膜。
N端序列分析
通过反相色谱在C1柱上分离His6-HdiR自切割反应的组分,使用乙腈在0.1%三氟乙酸中的线性梯度。在SMARTTM液相色谱仪上自动收集峰,并使用Procise 494 A测序仪进行N端序列分析。
体内HdiR稳定性的测定
通过将乳酸乳球菌菌株在30°C培养至OD600=0.5,然后加入3μM MMC,1分钟后加入CAM以阻断蛋白质合成,研究HdiR在MMC存在下的稳定性。在所示时间点取样,在离心前加入蛋白酶抑制剂混合物。通过将指数生长的KS78、KS79和KS80细胞从25°C转移到38.5°C,研究热应激条件下HdiR的稳定性。5或15分钟后用CAM阻断蛋白质合成,在所示时间取样。用玻璃珠匀浆细胞并离心后获得无细胞提取物。用蛋白质测定试剂盒测定蛋白质浓度。等量的蛋白质在SDS-PAGE中分离,并通过Western印迹分析HdiR的稳定性。
结果
从乳酸乳球菌乳脂亚种MG1363中鉴定HdiR
为了寻找乳酸乳球菌Clp蛋白酶的底物蛋白,我们检测了乳酸乳球菌乳脂亚种MG1363基因组是否编码UmuD的同源物。使用针对大肠杆菌UmuD的多克隆抗体进行的蛋白质印迹分析显示,乳酸乳球菌MG1363不编码UmuD的紧密同源物。此外,对乳酸乳球菌乳酸亚种IL1403基因组序列的搜索未能发现umuD基因,尽管遗传证据已确定乳酸乳球菌IL1403中存在UmuC。然而,当我们使用存在于肺炎链球菌基因组中的Tn5252转座子元件编码的UmuD样蛋白进行搜索时,我们检索到一个具有32%同一性的蛋白质。相似性最高的区域集中在LexA蛋白家族典型的三个保守结构域,包括LexA、UmuD、MucA、RumA和cl样噬菌体阻遏物。参与大肠杆菌LexA的RecA依赖性自切割的氨基酸Ala84、Gly85、Ser119和Lys156在IL1403 YnaB中也保守。与转座子UmuD不同,YnaB携带一个假定的N端螺旋-转角-螺旋基序,因此被归类为假定的转录调节因子。因此,基于序列分析,YnaB与LexA和UmuD都有相似之处。值得注意的是,针对IL1403基因组的BLAST搜索表明乳酸乳球菌缺乏保守的LexA。
为了进一步表征YnaB,我们对来自MG1363的YnaB直系同源基因进行了测序,并根据我们后续的研究,将其命名为热激和DNA损伤诱导调节因子。hdiR基因编码一个252个氨基酸残基的蛋白质,计算分子量为28.7 kDa。推导的HdiR氨基酸序列与IL1403的YnaB具有90%的同一性,并且在链球菌和葡萄球菌物种中存在同源物。虽然这些未表征的蛋白质与HdiR的总体氨基酸同一性仅为26-32%,但围绕LexA蛋白自切割典型残基的氨基酸是保守的,并且它们都包含一个N端HTH基序。HdiR同源物和LexA蛋白的多重比对作为补充材料提供。
在乳酸乳球菌MG1363中,hdiR基因前面有一个假定的保守原核营养启动子、一个核糖体结合位点以及两个反向重复。较远的反向重复2类似于典型的rho非依赖型转录终止子,可能终止上游定位的ynaC的表达。近端的反向重复1位于假定的-35区上游两个核苷酸处。在hdiR下游未发现明显的转录终止子结构,表明hdiR与下游定位的ynaA基因形成双顺反子操纵子。
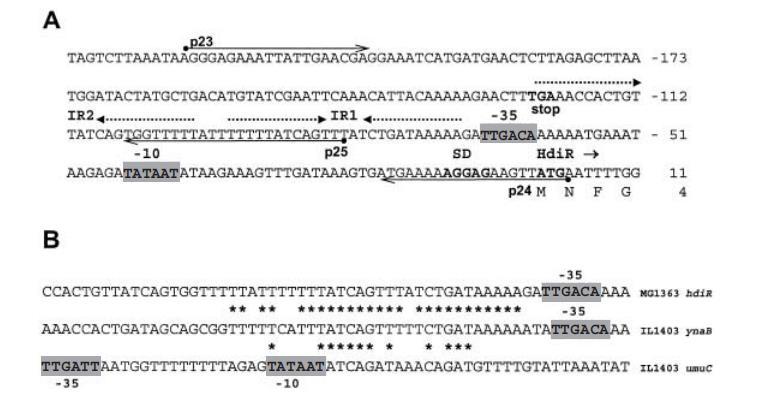
图1. MG1363 hdiR区域的部分核苷酸序列。A. 假定的调控区域和HdiR编码区5'端的DNA序列。预测的-10/-35六核苷酸用框标出。上游ynaC的翻译终止密码子、假定的核糖体结合位点和HdiR的起始密码子用粗体显示。IR1和IR2用虚线箭头表示。以点开始的箭头显示用于DNA结合实验生成DNA片段的引物。B. 围绕hdiR的IR1区域以及IL1403中ynaB和umuC基因前的IR1样序列的核苷酸序列。星号表示相同的核苷酸。
hdiR在乳酸乳球菌MG1363中受热和DNA损伤调控
在大肠杆菌中,LexA的自调控表达由DNA损伤条件诱导。由于HdiR类似于LexA,我们检测了DNA交联剂丝裂霉素C是否能诱导HdiR表达。对乳酸乳球菌MG1363的Northern印迹分析显示了一个1.0 kb的单一hdiR特异性转录本,表明hdiR与位于其下游的ynaA基因形成一个双顺反子操纵子。此外,MG1363中的hdiR表达在暴露于MMC 20分钟后被诱导约15倍。
我们还检测了hdiR是否被其他环境条件诱导,例如在38.5°C热激、在10°C冷激、低pH或氯化钠,并观察到只有热激影响hdiR表达。虽然在MMC存在下hdiR表达的最佳诱导在15到30分钟之间达到,但在将细胞转移到升高的温度后5分钟就已经达到约15倍的诱导。奇怪的是,在升高温度下长时间培养表明,hdiR表达在20分钟后就被重新抑制回诱导前的水平。然而,在MMC存在下,60分钟后hdiR表达水平仍是非诱导水平的六倍。
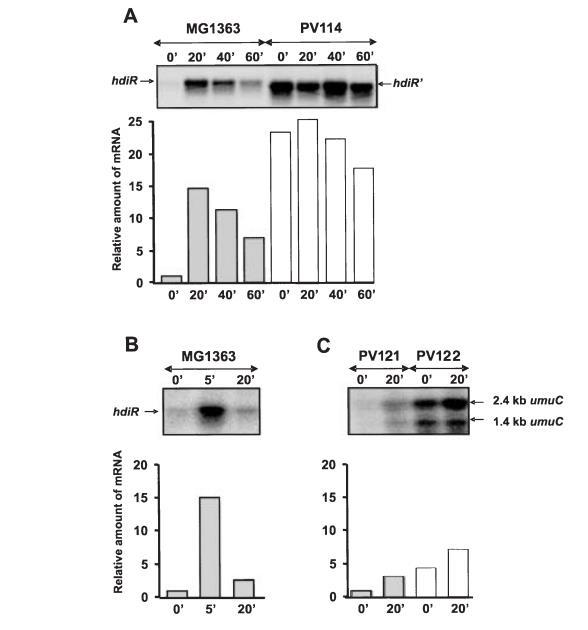
图2. hdiR和umuC基因表达的Northern分析。A. 添加MMC前和添加后20、40、60分钟,MG1363和PV114细胞中的hdiR表达。B. 从25°C转移到38.5°C前和转移后5、20分钟,MG1363细胞中的hdiR表达。C. 添加MMC前和添加后20分钟,PV121和PV122细胞中的umuC表达。柱状图显示相对mRNA诱导比率,通过将RNA样品的信号除以0分钟时MG1363细胞的信号计算得出。膜上的RNA量在rRNA杂交后进行校正。
相关新闻推荐
1、驴用微生态制剂干酪乳酸杆菌筛选、生理生化鉴定及生长条件(二)
2、UL56基因下游重组病毒对鸭肠炎病毒生物特性、生长曲线影响——材料与方法
3、伤寒沙门菌t4606基因缺陷株在高渗应激环境下的生长曲线(二)