乳杆菌的生长。
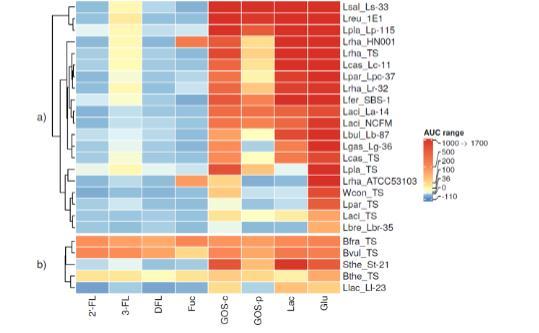
图3. 碳源对乳杆菌科及其他人体共生菌生长的影响。在1% 2'-岩藻糖基乳糖(2'-FL)、3-岩藻糖基乳糖(3-FL)、二岩藻糖基乳糖(DFL)、岩藻糖(Fuc)、商品化低聚半乳糖(GOS-c)、富集低聚糖的GOS(GOS-p)、乳糖(Lac)和葡萄糖(Glu)存在条件下,(a)乳杆菌科及(b)其他人體共生菌或食品相关细菌的生长曲线下面积(AUC)评估。通过减去未添加碳水化合物的生长培养基中细菌的AUC值对AUC值进行标准化。生长数据采用色标编码:深红色代表强烈生长(AUC 1000-1700;500-999;200-499;100-199;36-99;0-35),深蓝色代表生长弱于基础培养基(AUC-110至-1)。细菌菌株缩写参见表2。
| 细菌物种 | 菌株名称 | 菌株来源 | 模式菌株/益生菌 | 培养基 | 温度(°C) | 气氛 | 缩写 |
|---|---|---|---|---|---|---|---|
| 青春双歧杆菌 | DSM 20083 | DSMZ | 模式菌株 | 58 | 37 | 厌氧 | Bado_TS |
| 动物双歧杆菌乳亚种 | DSM 10140 | DSMZ | 模式菌株 | 58 | 37 | 厌氧 | Blac_TS |
| 动物双歧杆菌乳亚种 | Bi-07 | DGCC | 益生菌 | 58 | 37 | 厌氧 | Blac_Bi-07 |
| 动物双歧杆菌乳亚种 | Bl-04 | DGCC | 益生菌 | 58 | 37 | 厌氧 | Blac_Bl-04 |
| 动物双歧杆菌乳亚种 | B420 | DGCC | 益生菌 | 58 | 37 | 厌氧 | Blac_B420 |
| 动物双歧杆菌乳亚种 | HN019 | DGCC | 益生菌 | 58 | 37 | 厌氧 | Blac_HN019 |
| 两歧双歧杆菌 | DSM 20456 | DSMZ | 模式菌株 | 改良RCM | 37 | 厌氧 | Bbif_TS |
| 两歧双歧杆菌 | Bb-06 | DGCC | 益生菌 | 58 | 37 | 厌氧 | Bbif_Bb-06 |
| 短双歧杆菌 | DSM 20213 | DSMZ | 模式菌株 | 58 | 37 | 厌氧 | Bbre_TS |
| 短双歧杆菌 | Bb-03 | DGCC | 益生菌 | 58 | 37 | 厌氧 | Bbre_Bb-03 |
| 链状双歧杆菌 | DSM 16992 | DSMZ | 模式菌株 | 58 | 37 | 厌氧 | Bcat_TS |
| 长双歧杆菌长亚种 | DSM 20219 | DSMZ | 模式菌株 | 58 | 37 | 厌氧 | Blon_TS |
| 长双歧杆菌长亚种 | Bl-05 | DGCC | 益生菌 | 58 | 37 | 厌氧 | Blon_Bl-05 |
| 长双歧杆菌婴儿亚种 | DSM 20088 | DSMZ | 模式菌株 | 58 | 37 | 厌氧 | Binf_TS |
| 长双歧杆菌婴儿亚种 | Bi-26 | DGCC | 益生菌 | 58 | 37 | 厌氧 | Binf_Bi-26 |
| 假小链双歧杆菌 | DSM 20438 | DSMZ | 模式菌株 | 58 | 37 | 厌氧 | Bpse_TS |
| 干酪乳杆菌 | DSM 20011 | DSMZ | 模式菌株 | MRS | 30 | 好氧 | Lcas_TS |
| 干酪乳杆菌 | Lc-11 | DGCC | 益生菌 | MRS | 30 | 好氧 | Lcas_Lc-11 |
| 副干酪乳杆菌 | DSM 5622 | DSMZ | 模式菌株 | MRS | 37 | 厌氧 | Lpar_TS |
| 副干酪乳杆菌 | Lpc-37 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Lpar_Lpc-37 |
| 鼠李糖乳杆菌 | DSM 20021 | DSMZ | 模式菌株 | MRS | 37 | 厌氧 | Lrha_TS |
| 鼠李糖乳杆菌 | HN001 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Lrha_HN001 |
| 鼠李糖乳杆菌 | ATCC 53103, GG | DGCC | 益生菌 | MRS | 37 | 厌氧 | Lrha_ATCC53103 |
| 鼠李糖乳杆菌 | Lr-32 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Lrha_Lr-32 |
| 植物乳杆菌 | DSM 20174 | DSMZ | 模式菌株 | MRS | 37 | 厌氧 | Lpla_TS |
| 植物乳杆菌 | Lp-115 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Lpla_Lp-115 |
| 嗜酸乳杆菌 | DSM 20079 | DSMZ | 模式菌株 | MRS | 37 | 厌氧 | Laci_TS |
| 嗜酸乳杆菌 | NCFM | DGCC | 益生菌 | MRS | 37 | 厌氧 | Laci_NCFM |
| 嗜酸乳杆菌 | La-14 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Laci_La-14 |
| 德氏乳杆菌保加利亚亚种 | Lb-87 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Lbul_Lb-87 |
| 加氏乳杆菌 | Lg-36 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Lgas_Lg-36 |
| 短乳杆菌 | Lbr-35 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Lbre_Lbr-35 |
| 唾液乳杆菌 | Ls-33 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Lsal_Ls-33 |
| 发酵粘液乳杆菌 | SBS-1 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Lfer_SBS-1 |
| 罗伊氏粘液乳杆菌 | 1E1 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Lreu_1E1 |
| 融合魏斯氏菌 | DSM 20196 | DSMZ | 模式菌株 | MRS | 30 | 好氧 | Wcon_TS |
| 脆弱拟杆菌 | DSM 2151 | DSMZ | 模式菌株 | PYG | 37 | 厌氧 | Bfra_TS |
| 多形拟杆菌 | DSM 2079 | DSMZ | 模式菌株 | PYG | 37 | 厌氧 | Bthe_TS |
| 普通拟杆菌 | DSM 1447 | DSMZ | 模式菌株 | PYG | 37 | 厌氧 | Bvul_TS |
| 乳酸乳球菌乳亚种 | Ll-23 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Llac_Ll-23 |
| 唾液链球菌嗜热亚种 | St-21 | DGCC | 益生菌 | MRS | 37 | 厌氧 | Sthe_St-21 |
| 艰难梭菌 | DSM 1296 | DSMZ | 模式菌株 | 改良RCM | 37 | 厌氧 | Cdif_TS |
| 艰难梭菌 | CCUG 46468 | CCUG | - | 改良RCM | 37 | 厌氧 | Cdif_CCUG46468 |
| 艰难梭菌 | NCTC 13574 | NCTC | - | 改良RCM | 37 | 厌氧 | Cdif_NCTC13574 |
| 产气荚膜梭菌 | E-98861 T | VTTCC | 模式菌株 | 改良RCM | 37 | 厌氧 | Cper_TS |
| 阪崎克罗诺杆菌 | DSM 4485 | DSMZ | 模式菌株 | TSB | 30 | 好氧 | Csak_TS |
| 阪崎克罗诺杆菌 | BAA-894 | ATCC | - | TSB | 37 | 好氧 | Csak_BAA-894 |
| 大肠杆菌 O1:K1:H7 | E94564T | VTTCC | 模式菌株 | TSB | 37 | 好氧 | Ecol_TS |
| 大肠杆菌 O111:K58(B4):H2 | CCUG 42878 | CCUG | - | TSB | 37 | 好氧 | Ecol_CCUG42878 |
| 大肠杆菌 O142:K86(B):H6 | CCUG 11442 | CCUG | - | TSB | 37 | 好氧 | Ecol_CCUG11442 |
| 大肠杆菌 O111:K58(B4):H- | CCUG 11412 | CCUG | - | TSB | 37 | 好氧 | Ecol_CCUG11412 |
| 大肠杆菌 O20:K84:H26 | NCTC 10865 | NCTC | - | TSB | 37 | 好氧 | Ecol_NCTC10865 |
| 肠道沙门氏菌肠道亚种;血清型鼠伤寒 | DSM 17058 | DSMZ | 模式菌株 | TSB | 37 | 好氧 | Sent_TS |
| 肠道沙门氏菌肠道亚种;血清型波那 | NCTC 4840 | NCTC | - | TSB | 37 | 好氧 | Sent_NCTC4840 |
| 肠道沙门氏菌肠道亚种;血清型鼠伤寒 | NCTC 13954 | NCTC | - | TSB | 37 | 好氧 | Sent_NCTC13954 |
| 福氏志贺氏菌 | DSM 4782 | DSMZ | 模式菌株 | TSB | 37 | 好氧 | Sfle_TS |
| 福氏志贺氏菌 | CCUG 39080 | CCUG | - | TSB | 37 | 好氧 | Sfle_CCUG39080 |
注:T=温度,单位为摄氏度;*预培养条件;58=改良DSMZ双歧杆菌培养基58;PYG=蛋白胨、酵母提取物、葡萄糖培养基;MRS=De Man、Rogosa和Sharpe培养基;改良RCM=改良强化梭菌培养基;TSB=胰蛋白酶大豆肉汤;ATCC=美国模式培养物保藏中心;DSMZ=德国微生物和细胞培养物保藏中心;DGCC=杜邦培养物保藏中心;VTTCC=VTT技术研究中心芬兰培养物保藏中心;NCTC=英国公共卫生署国家模式培养物保藏中心;CCUG=哥德堡大学培养物保藏中心。
乳杆菌科家族的碳水化合物利用以热图形式显示在图3a中。从每种碳源的AUC中,减去了未添加碳水化合物的培养基的AUC。然后根据生长模式的相似性对乳杆菌科细菌菌株进行聚类,形成了三个主要组。第一个簇在两种GOS产品、乳糖和葡萄糖上均表现出剧烈生长,仅包括唾液乳杆菌Ls-33、罗伊氏粘液乳杆菌1E1(Lreu_1E1)和植物乳杆菌Lp-115(Lpla_Lp-115)。影响其他两个簇(图3a)聚类的主要因素是无法利用乳糖(最下面的簇,6种细菌)和3-FL与GOS-p利用的差异(第二个簇,11种细菌)。
19种来自乳杆菌科的细菌均未在2'-FL或DFL上生长(图3a)。正如Yu等人先前所示,鼠李糖乳杆菌GG(Lrha_ATCC53103)在我们的研究中也未在2'-FL、3-FL或DFL上生长。然而,我们观察到其余三种研究的鼠李糖乳杆菌菌株(Lrha_TS、Lrha_HN001和Lrha_Lr-32)在3-FL上有轻微生长,这可能是由于乳糖利用的差异。3-FL也支持干酪乳杆菌(Lcas_Lc-11)、植物乳杆菌、发酵粘液乳杆菌(Lfer_SBS-1)、唾液乳杆菌和罗伊氏粘液乳杆菌(Lreu_1E1)的轻微生长。先前已显示干酪乳杆菌BH23可以在岩藻糖基-1,3-N-乙酰葡糖胺上生长。然而,此处研究的干酪乳杆菌菌株(Lcas_TS和Lcas_Lc-11)未在2'-FL和DFL上生长,仅观察到3-FL上的弱生长。
鼠李糖乳杆菌菌株GG(Lrha_ATCC53103)和HN001(Lrha_HN001)是本研究中乳杆菌科中唯一在岩藻糖上生长良好的细菌。Thongaram等人也报道了类似的发现。然而,这种现象在所有研究的鼠李糖乳杆菌菌株中并不一致,因为其他两种鼠李糖乳杆菌菌株(Lrha_TS和Lrha_Lr-32)未在岩藻糖上生长。
先前在乳杆菌对两种GOS产品及其商业和纯化版本的利用中发现了物种和菌株特异性差异。根据早期的一项研究,罗伊氏粘液乳杆菌似乎能有效利用GOS、葡萄糖和乳糖。在当前研究中,罗伊氏粘液乳杆菌(Lreu_1E1)在两种GOS产品和葡萄糖上也生长良好。此处研究的大多数乳杆菌科菌株无法像利用商业GOS那样好地利用纯化GOS。嗜酸乳杆菌和副干酪乳杆菌的模式菌株(分别为Laci_TS和Lpar_TS)在GOS或乳糖上仅微弱生长或根本不生长,而益生菌菌株嗜酸乳杆菌La-14(Laci_La-14)、嗜酸乳杆菌NCFM和副干酪乳杆菌Lpc-37(Lpar_Lpc-37)在GOS和乳糖上生长良好,表明嗜酸乳杆菌和副干酪乳杆菌物种在GOS和乳糖利用方面存在菌株依赖性变异。
已知嗜酸乳杆菌NCFM表达LacS作为将GOS和乳糖转运到细菌细胞中的唯一转运蛋白,这可能解释了NCFM与模式菌株在GOS和乳糖利用方面的差异。早期,另外两种嗜酸乳杆菌菌株和嗜酸乳杆菌NCFM在三种不同GOS产品上生长良好,而在另一项研究中,三种不同嗜酸乳杆菌菌株在GOS利用方面存在差异。植物乳杆菌模式菌株未在乳糖上生长,而植物乳杆菌Lp-115(Lpla_Lp-115)在乳糖上生长良好。融合魏斯氏菌模式菌株(Wcon_TS)仅利用葡萄糖和商业GOS。
除了证实先前使用HMOs和GOS的生长研究外,我们的结果突出了碳水化合物利用的菌株特异性差异。这些结果证明了物种内部甚至同一物种菌株之间在碳源利用和生长强度方面的差异。
拟杆菌和乳杆菌目的生长。
我们的研究还包括了一些人类共生菌的模式菌株。它们的碳水化合物利用结果以热图形式显示在图3b中。在本研究中,脆弱拟杆菌(Bfra_TS)、普通拟杆菌(Bvul_TS)和多形拟杆菌(Bthe_TS)的模式菌株在HMOs 2'-FL、3-FL和DFL上生长良好,与在葡萄糖和乳糖上观察到的生长相似,这与先前的研究一致。类似地,我们发现所有研究的拟杆菌菌株在商业和纯化GOS上都生长良好,这也与先前对多形拟杆菌、脆弱拟杆菌和普通拟杆菌的研究一致。
相反,一些先前的研究表明,在临床环境中,GOS消耗导致拟杆菌属丰度下降,并且在体外模拟实验中,通过微阵列评估,细胞数量减少。多形拟杆菌以其能够利用多个多糖利用位点在许多复杂的宿主和膳食碳源上良好生长而闻名。这与我们的发现一致,即多形拟杆菌在所有碳源上都生长良好,并且不同碳源之间的生长变化相对较小。碳源有助于各种PULs的表达。多形拟杆菌在GOS上的生长激活了与粘蛋白聚糖和半乳聚糖利用相关的PULs,包括糖苷水解酶GH53(半乳聚糖酶),该酶在细胞外将较长的GOS分子降解为较小的分子。先前已显示拟杆菌属在HMOs上的生长诱导了与粘蛋白利用相关的PULs。与双歧杆菌相反,所有评估的拟杆菌属菌株都能够利用岩藻糖。
该组中的另外两种细菌物种,乳酸乳球菌乳亚种Ll-23(Llac_Ll-23)和嗜热链球菌St-21(Sthe_St-21),属于乳杆菌目,因此与其他乳杆菌具有共同特征。它们未在测试的HMOs或岩藻糖上生长。嗜热链球菌St-21(Sthe_St-21)在葡萄糖、乳糖和GOS-c上生长良好,尽管在GOS-p上生长减少。先前一项关于嗜热链球菌FUA3194的研究也显示其水解GOS,但不水解HMOs。乳酸乳球菌Ll-23(Llac_Ll-23)未在GOS-p上生长,仅在葡萄糖、乳糖和GOS-c上生长。类似地,先前一项研究发现20种乳酸球菌菌株中均未在galactosyl lactose上生长且无β-半乳糖苷酶活性。
相关新闻推荐
1、明永冰川地区低温黄杆菌噬菌体生长曲线绘制及生物学特性(四)
2、450 MPa高压联合乳过氧化物酶系统对冷熏鲑鱼中食源性病原体的杀菌动力学
3、禽呼肠病毒S1133毒株接种DF1细胞一步生长曲线的绘制——结果、结论