2.材料与方法
2.1.细菌菌株、噬菌体扩增和生长条件
噬菌体φR1-37按照先前描述的方法[16]在小肠结肠炎耶尔森菌O:3菌株YeO3-R1[15]中扩增。小肠结肠炎耶尔森菌在室温下于溶菌肉汤(LB)中培养。所有固体培养物使用Luria琼脂(LA)平板,通过在LB中添加1.5%的Bacto琼脂制备。
2.2.生长曲线
将过夜(16小时)细菌培养物用新鲜LB培养基按1:10稀释,将180μL等分试样分配到蜂蜜comb板孔中(Growth Curves Ab Ltd.,Helsinki,Finland),并与20μL的φR1-37噬菌体储备液(1 x 10^9 PFU/mL)的10^-2至10^-6稀释液等分试样混合。因此,近似感染复数(MOI)范围从10^-1到10^-5。在阳性对照培养物中,噬菌体被20μL培养基替代,阴性对照通过将20μL噬菌体储备液与180μL培养基混合制备。生长实验在4°C、16°C、22°C和37°C下使用Bioscreen C培养箱(Growth Curves Ab Ltd.)在连续振荡下进行。在选定的时间间隔测量光密度(OD)OD600值。平均值根据在八个平行孔中生长的细菌获得的值计算。
2.3.总RNA提取
用于转录组分析时,将YeO3-R1细菌在室温(22°C)下培养16小时,随后用新鲜LB按1:10稀释至总体积10 mL。在培养期间测量培养物的OD600,当达到0.6时,将培养物分成两个相同的部分。一部分用MOI为10的φR1-37噬菌体感染,另一部分作为未感染对照。接下来,两个培养样品在相同条件下培养,并测量培养物的OD600以确保感染过程正常。在感染后不同时间点(2、5、10、15、21、28、35、42、49分钟)从非感染培养物(阴性对照)和感染培养物中取样品(1 mL)进行RNA分离。使用SV总RNA分离系统(Promega,Madison,Wisconsin,US)从样品中分离总RNA。使用Bioanalyzer(Agilent,Santa Clara,California,US)和RNA 6000 Nano Kit(Agilent)分析分离的RNA制备物的质量和核糖体RNA(rRNA)谱。
2.4.RNA测序
RNA测序和数据分析在芬兰分子医学研究所(FIMM)技术中心测序单元进行。使用针对革兰氏阴性菌的Ribo-ZeroTM rRNA去除试剂盒(Epicentre,Madison,Wisconsin,US)去除rRNA。在Illumina HISeq2000测序仪(Illumina,San Diego,California,US)上进行双末端测序,读长为90个核苷酸。RNA序列数据已存入Gene Expression Omnibus(登录号GSE77068)。
2.5.计算和统计分析
原始测序读数在进行质量修剪和过滤后,使用CLC genomics workbench v7.5.1映射到宿主和噬菌体基因组。计算比对到噬菌体和宿主基因组中每个非rRNA基因特征的总基因读数(TGR)。
如果计算出的TGR折叠变化(log2FC)值的log2值超过1.5或低于-1.5,并且阴性对照值小于样本平均计数值减去2个标准差或高于样本平均计数值加上2个标准差,则认为基因差异表达。在比较早期和晚期细菌反应时,使用Student t检验测试差异表达。如果p<0.01,则认为差异表达具有统计学意义。那些包含0分钟阴性对照的比较确实容易受到显著培养偏差的影响,因为该条件只有一个重复。
通过Total Count[23]标准化后,对φR1-37时间模式进行聚类如下。计算2-5分钟(早期)、10-21分钟(中期)、28-49分钟(晚期)TGR值的平均值,并以百分比值表示,最高值设为100%。根据最高平均值对应的阶段对基因进行分类。如果不同阶段之间的平均值差异小于40%,则认为基因未被调控。平均TGR<10的基因不再进一步考虑。
3.结果与讨论
3.1.φR1-37感染过程的微生物学参数
先前已使用一步生长曲线法确定了噬菌体φR1-37感染的动力学,显示潜伏期为40分钟,裂解期为50分钟[16]。在此,我们使用Bioscreen C进一步分析了不同温度条件下的噬菌体-宿主相互作用。结果表明,φR1-37在所有研究的温度下都能在YeO3-R1中建立感染,但其有效性随温度升高而增加(图1)。在4°C和10°C时,只有最高数量的噬菌体(即MOI=100)能够影响细菌生长。在10°C时,培养物被跟踪足够长的时间,可以观察到细菌在40至70小时孵育期间缓慢裂解(图1)。
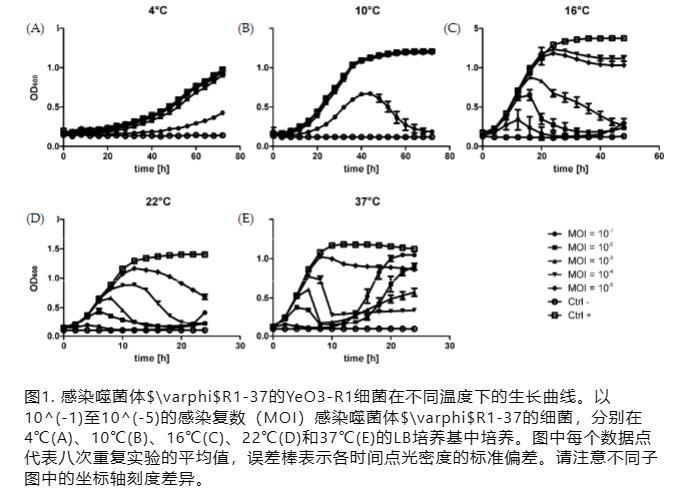
在较高温度下,使用所有其他MOI值也进行了感染,细菌生长曲线很好地反映了MOI的范围。在16°C、22°C和37°C下,分别在(完全)裂解的培养物中于40、20和15小时后检测到细菌的再生长。该实验证明噬菌体φR1-37能够在较宽的温度范围内建立有效感染。在较高温度下完全裂解培养物后出现的再生长很可能是由于出现了噬菌体抗性细菌。由于φR1-37的受体是LPS的OC,噬菌体抗性自发突变体以高频率出现[16]。基于这些结果,用于RNA测序分析的样品从在室温下培养并以MOI=10感染的细菌中收集。
相关新闻推荐
1、可降胆固醇的犬源植物乳杆菌分离与鉴定、生长曲线、益生性和安全性研究(一)
3、CSP-MDA-7/IL-24重组蛋白肝靶向肽-抗肿瘤肽的最佳表达条件(二)