经过24小时培养期后,API 50CH测试证实了IL1403△claR完全无法发酵乳糖和纤维二糖。虽然IL1403△claR和IL1403△ccpA△claR突变体无法同化乳糖的情况持续了36小时,但在最初的24小时后,在IL1403△claR中观察到了纤维二糖的有效利用(图2d)。当通过API 50CH评估时,在IL1403△ccpA△claR与其亲本IL1403△ccpA之间在纤维二糖发酵方面未发现差异(图2d)。
使用表型微阵列方法的全局代谢活性测量表明,缺失claR导致IL1403△claR中乳糖利用的完全废除。相反,当纤维二糖作为唯一碳源可用时,与野生型相比,这仅导致IL1403△claR代谢活性的轻微下降(数据未显示)。
在API 50CH上可用的所有其他47种糖或Biolog PM1和PM2阵列上可用的188种碳源上,未发现测试菌株之间进一步的显著差异表型。
将pIL253claR转入IL1403△claR,产生IL1403△claRpIL253claR菌株,完全互补了claR缺失的影响,恢复了突变体在含纤维二糖和/或乳糖的培养基中的生长(数据未显示)。
ClaR以低水平组成型表达且独立于CcpA介导的调控 除了由CcpA结合cre序列介导的直接基因表达调控外,先前也观察到间接控制。尽管claR上游DNA区域缺乏类似于cre框的序列,但其表达仍可能以间接方式受CcpA调控。为了测试这种现象是否也发生在claR的调控中,以及其表达是否受不同糖存在的影响,通过实时定量RT-PCR估计了在IL1403和IL1403△ccpA中响应熊果苷、纤维二糖、半乳糖和葡萄糖的claR表达水平。在这两种菌株中以及所有测试的糖存在下,claR基因均以低水平组成型表达,达到大约0.05的平均相对基因表达水平值(数据未显示)。这表明claR的表达不受任何测试糖的诱导或抑制,并且不经历CcpA依赖性调控。
celB、bglS和ptcBA基因的表达在纤维二糖存在下受到ClaR的正向调控 先前,我们显示编码PTSCel-Lac组分(celB和ptcBA)和bglS的基因在乳酸乳球菌IL1403中受到CcpA的负调控。为了估计ClaR对不同碳源存在下bglS、celB和ptcBA表达的影响,从在C-CDM、G-CDM、Gal-CDM或GalC-CDM中生长的IL1403和IL1403△claR的独立细菌培养物中提取RNA。GalC-CDM中的半乳糖用于支持IL1403△claR的生长,因为该突变体无法利用纤维二糖作为碳源。由于乳酸乳球菌IL1403与其他使用的糖(如纤维二糖、半乳糖或葡萄糖)相比生长参数(生长速率、延滞期和最终培养物光密度)较差,未将乳糖纳入糖阵列中。
在Gal-CDM中,与G-CDM相比,野生型IL1403和IL1403△claR菌株中bglS、celB和ptcBA的表达水平仅略有变化,而ptcBA的转录在两种菌株中在补充半乳糖的培养基中均增加(表2)。IL1403野生型和IL1403△claR菌株在半乳糖或葡萄糖存在下可比较的表达水平(ClaR激活比率~1;表2)意味着这些糖不会引起bglS、celB和ptcBA的ClaR依赖性激活。
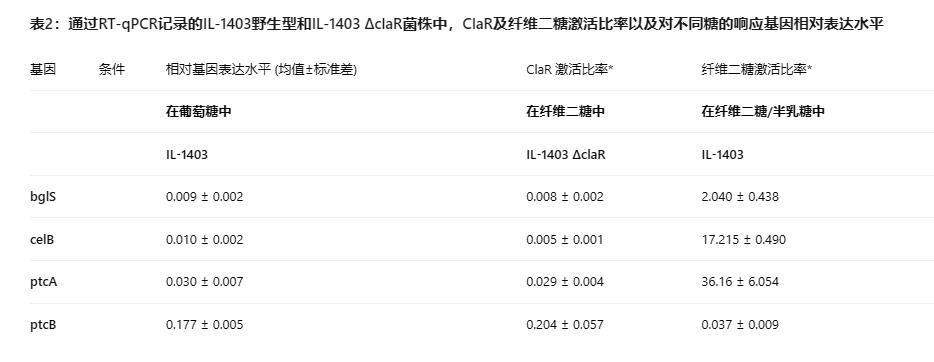
当菌株在GalC-CDM中生长时,IL1403野生型和IL1403△claR之间bglS和celB表达水平的显著差异是明显的。在这种情况下,ClaR存在下(在IL1403中)celB和bglS的表达水平显著高于ClaR缺失下(在IL1403△claR中)获得的值,分别达到8和36的ClaR激活比率(计算为IL1403和IL1403△claR中相对基因表达的商)(表2)。对于ptcBA,ClaR激活比率接近3,表明ClaR依赖性调控较小(表2)。
当野生型菌株在补充纤维二糖(C-CDM)作为唯一碳源的培养基中生长时,检测到所有测试基因的转录水平显著更高(表2)。添加半乳糖(GalC-CDM)导致与C-CDM相比,bglS、celB和ptcAB的转录下降2-4倍。这可能表明半乳糖的存在以类似于葡萄糖的方式导致转录抑制。
此外,为了确定纤维二糖对基因表达的影响,计算了细胞二糖激活比率,作为在GalC-CDM和Gal-CDM中生长的野生型菌株IL1403或IL1403△claR中相对基因表达的商。在IL1403中获得的这些比率从25(bglS)和7(celB)到低至3-4倍(ptcBA)不等,意味着纤维二糖对相关基因的激活有不同的影响水平(表2)。这些纤维二糖激活比率与为GalC-CDM计算的ClaR激活值相似,表明这种纤维二糖依赖性调控可能是由于ClaR的作用。这通过观察在ClaR缺失下获得的纤维二糖激活比率通常围绕因子1得到证实,表明即使在纤维二糖存在下,IL1403△claR中也缺乏基因表达调控(表2)。