3.核心稀有类群的持久性的生态驱动因素
如前所述,核心微生物组由特定环境所特有的分类群、相关基因和功能属性组成。尽管多年来定义有所不同,但核心微生物组通常被认为是由那些在该环境中普遍存在且稳定的分类群所构成的。在空间和时间上始终存在于环境中的微生物群落;也就是说,它们在数量上具有很高的占有率和稳定性。它们对于维持肠道和口腔健康以及增强微生物群落对环境压力的抵抗力具有重要意义,因此引发了大量研究。相比之下,在撰写本文时,我们尚未发现有关核心稀有微生物群的研究。由于丰度占有率和时间动态对于确定核心微生物组成员至关重要,因此使用相同的参数来确定核心稀有微生物群成员是合理的。有趣的是,最近的一项研究调查了亚热带水库不同深度和时间点核心微生物群和卫星微生物群(类似于稀有微生物群)的时空变化。
该研究发现,稀有微生物群与核心微生物群在时空维度上的存在具有一致性。尽管他们没有专门研究核心稀有微生物群,但有可能永久性稀有微生物群的成员在时间尺度上以低丰度存在于核心微生物群中。然而,目前尚无研究对这一假设进行实证验证,因此,导致核心稀有物种持续存在的因素也尚未得到探索。在接下来的部分中,我们将尝试确定能够支持核心稀有物种持续存在的生态和基因组驱动因素(图2)。
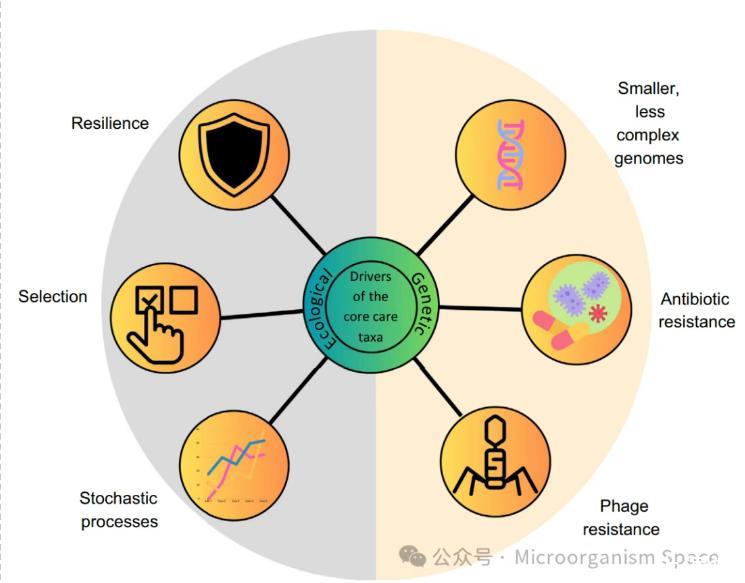
图2|核心稀有类群持久存在的驱动因素。生态(恢复力、选择、随机过程)和遗传(基因组的性质、存在赋予噬菌体和抗生素抗性的基因岛)驱动核心稀有类群持久存在的因素。核心稀有类群的恢复力和环境选择可作为生态驱动因素,在其持久存在中发挥重要作用。此外,稀有生物群落的形成已被发现显著受到诸如遗传漂变和随机生态相互作用等随机过程的影响。此外,较小的基因组规模和较低的复杂性可能是核心稀有类群在所栖息的狭窄生态位中持久存在的重要遗传决定因素。核心稀有类群成员中存在的基因岛可以编码抗生素抗性基因,这可能进一步帮助其中一些生物与丰富类群竞争并持久存在。
3.1对时空异质性的抗性
微生物对环境的响应因代谢能力的抗性或恢复力而异。由于时空异质性导致的环境波动可能会使生物体面临权衡取舍,从而失去某些特性而保留其他特性。支持这一观点的证据可以在诸如YAS框架等生命策略框架中找到,该框架指出生物体要么是高生物量的高产型(Y),要么是资源获取专家型(A),要么是耐逆境型(S),但不可能同时具备这三种类型。同样,Grime提出的CSR框架指出,物种可以是竞争者(C)、耐逆境者(S)或先锋物种(R),这取决于它们所处的环境和所具有的特性。在接下来的段落中,我们将尝试运用这些生态学框架来解释核心稀有、核心优势和非核心类群在应对环境时空波动时可能表现出的不同形式的恢复力。核心稀有生物群落是微生物群落中遗传和功能相似性的储存库,对群落的多样性和功能冗余性有显著贡献。文献中有若干报告指出稀有物种对环境波动具有显著的抗性;然而,驱动这种抗性的因素尚不明确。在特定选择压力下,种群中的某些个体可能会表现出有利的性状,因此抗性可能是固有的,由遗传倾向决定。此外,种群可以在选择压力下获得性状,这些进化而来的性状可以传递给后代。
与免疫系统相类比,我们可以将抗性大致分为两类:先天抗性,源于遗传或代谢库;适应性抗性,由生物或非生物因素造成的选择压力所导致的适应而产生。一代中的适应性抗性有可能在下一代中表现为先天抗性。每一种类型的韧性都由众多因素驱动(见图3)。微生物组可以被视为由三个部分组成:优势核心成员、核心稀有类群和非核心成员。优势核心成员具有高丰度和高占有率的特点,核心稀有类群则表现为低丰度和高占有率,而非核心成员则丰度多变且占有率低(图1)。有趣的是,将这些提出的丰度-占有率特征与YAS和CSR框架相结合,使我们能够对核心稀有类群、优势核心成员和非核心成员进行分类,并预测先天和适应性恢复力如何对其持久性产生不同的影响。例如,由于核心稀有类群具有高占有率和高度适应性恢复力,它们可以被归类为压力耐受者(S),因为它们能够在低丰度的情况下适应和进化以应对环境干扰。同样,优势核心成员可以被视为高效竞争者(C/Y),其丰度-占有率模式更多地受到先天恢复力的影响。最后,非核心成员由于丰度多变且占有率低,可能被归类为资源收集者(A/R)。由于它们的生态位因时空异质性而受到干扰,它们可能会利用诸如细胞外酶介导的资源解聚等机制来获取养分。然而,很难准确预测先天和适应性恢复力对丰富度-占有率的贡献,因此未来的研究有必要通过机制实验来探究这些模式。
3.2确定性和随机过程的影响
与任何生物物种一样,核心稀有物种在时间上的持续存在会受到确定性因素(生态位竞争、扩散过滤、选择)和随机因素(出生、死亡)。例如,环境选择可以作为核心稀有分类群持续存在的生态驱动因素,影响其丰度和分布。选择压力可分为同质性(由相似的环境条件驱动)或异质性(由多种不同的环境条件驱动)。通常,同质性选择压力会导致样本间的组成差异较小,而异质性选择会因分类群之间的适应性差异而增加差异。核心稀有微生物群可能以类似的方式被选择。例如,来自盐碱环境的样本可能包含耐盐的核心稀有分类群,而在异质性选择下的样本则会包含混合种群。同质性选择已知会对稀有生物群的组合产生显著影响。由于核心稀有分类群可能具有较高的分布率(在多个样本中存在,即样本间的组成差异较小),因此这些分类群也可能受到同质性选择的显著影响。
然而,在缺乏足够的经验数据的情况下,很难准确预测微生物组对同质选择和异质选择的反应。总的来说,似乎有理由假设,无论核心微生物组的稀有程度或丰富程度如何,其形成在一定程度上是由类似的过程所塑造的,这些过程有助于其在时空上的持续存在。除了确定性过程外,随机过程也会影响核心稀有类群的持久性。这些核心稀有种群数量少(丰度低),但分布范围广且对栖息地的特异性低(占有率高),这意味着它们在样本中普遍存在。尽管文献中有人认为诸如生态漂变之类的随机过程会影响一般稀有种群,但核心稀有种群的高占有率表明,它们可能更多地受到确定性过程(如选择)的影响,而非随机性的影响。然而,这并不一定意味着早期的理论不再成立,因为随机过程可能在塑造其他类型稀有种群的组成和动态方面仍发挥着重要作用。
3.3躲避捕食微生物群落的形成
受到自上而下(捕食)和自下而上(营养限制、环境条件等)驱动因素的共同影响。噬菌体是细菌最强大的天然捕食者,对微生物群落动态有着显著影响。值得注意的是,它们通过负频率依赖选择,专门针对群落中的优势菌群进行捕食,即噬菌体捕食那些数量最多的细菌菌群。这种捕食方式被称为“杀胜者”,它能确保单一菌群不会占据主导地位,有助于维持群落多样性。这也意味着数量较少的微生物菌群可以逃脱捕食,因为它们遭遇捕食性噬菌体的机会更少。
事实上,多项针对肠道微生物群的研究发现,在患病状态下,优势致病菌的数量会大幅减少,而在噬菌体疗法之后,通常被致病菌排挤的有益菌群会得以恢复。鉴于此类证据,核心稀有类群由于其丰度低而可避免噬菌体捕食,而核心优势类群则易受捕食。对于非核心类群,捕食的影响可能取决于具体情境,因为该组成员的丰度可能变化不定。然而,探究其机制至关重要。核心稀有物种如何维持其时间上的持久性以及噬菌体捕食在塑造其持久性方面所起的作用。
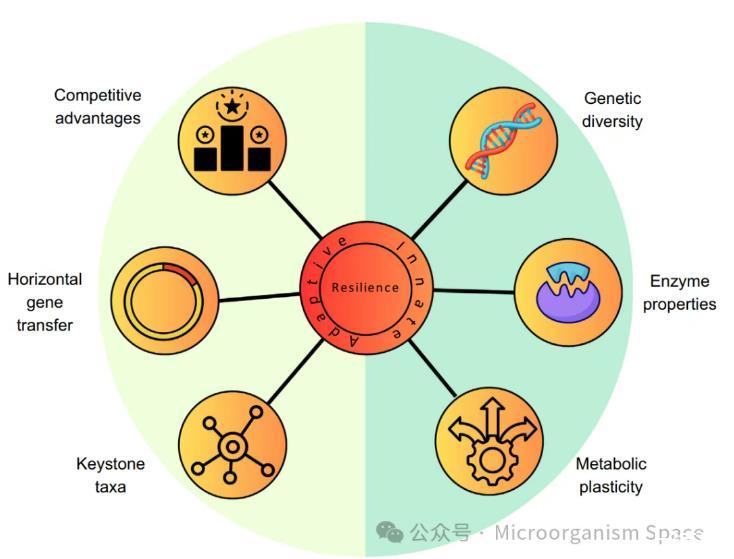
图3|核心稀有类群的恢复力驱动因素。核心稀有类群的内在(遗传多样性、酶特性、代谢可塑性)和适应性(竞争优势、水平基因转移、与关键类群的关联)恢复力的驱动因素。核心稀有类群的高遗传多样性,以及具有高底物亲和力和高代谢可塑性的酶,能够使这些类群高效利用其生态位中的稀缺资源,从而有助于其“内在”恢复力。此外,在持续环境压力下进化所获得的有利适应性可以使核心稀有类群获得竞争优势并得以持续存在。“适应性”恢复力也可能通过水平基因转移获得的抗性基因以及与群落中的关键类群的关联而获得。
相关新闻推荐
1、不同氮源、无机盐、ph、温度对多形炭角菌生长的影响(一)