2.3噬菌体PJNB028系统发育进化分析
基于全基因组序列的系统发育进化树显示(图3),与噬菌体PJNB028关系最近的是枯草芽孢杆菌噬菌体2S-4,这16株噬菌体均属于Caudoviricetes纲,其中3株属于Tsamsavirus属、1株属于Haetaevirus属、2株属于Spbetavirus属、1株属于Zhangjivirus属、9株未分类。基于末端大亚基蛋白质序列系统发育树显示(图4),与噬菌体PJNB028关系最近的是枯草芽孢杆菌噬菌体PK-3,13株噬菌体均属于Caudoviricetes纲,其中2株属于Tsamsavirus属、1株属于Natevirus属、2株属于Spbetavirus属、1株属于Kirovvirus属、1株属于Haetaevirus属、1株属于Kenyattavirus属、5株未分类。
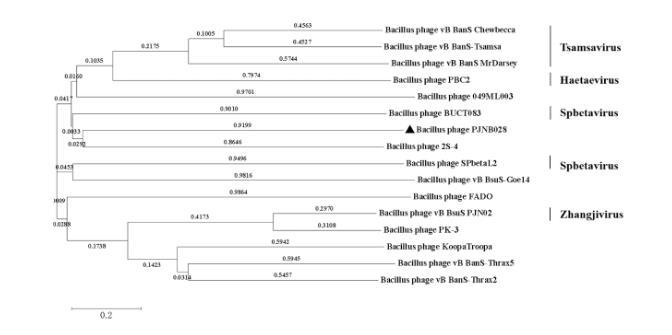
图3噬菌体PJNB028全基因组进化树
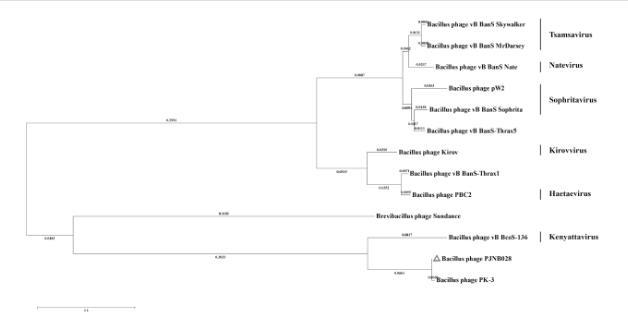
图4噬菌体PJNB028末端酶大亚基系统发育树
2.4噬菌体PJNB028最佳MOI测定
如图5所示,MOI为1时,噬菌体PJNB028的效价最高,达到4.5×108 PFU/mL。
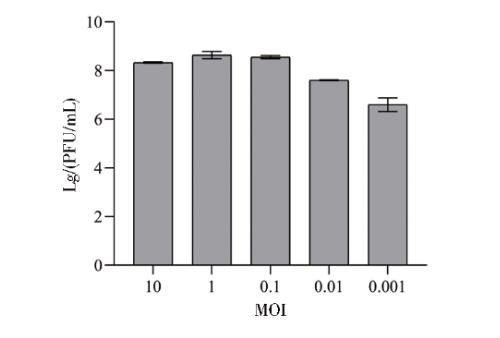
图5噬菌体PJNB028最佳MOI测定
2.5噬菌体PJNB028一步生长曲线
如图6所示,噬菌体PJNB028潜伏期为10 min,10~80 min效价开始明显上升,裂解期为60 min。爆发量为291 PFU/cell。
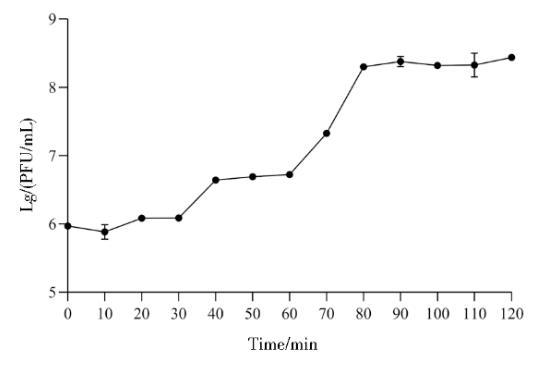
图6噬菌体PJNB028的一步生长曲线
2.6噬菌体PJNB028的温度稳定性测定
噬菌体在37~60℃环境下效价稳定。在70℃下孵育30 min,噬菌体效价由4×109 PFU/mL降至6.7×107 PFU/mL,孵育60 min降至8.17×106 PFU/mL;在80℃下孵育30 min,噬菌体效价从4×109 PFU/mL降至6.1×104 PFU/mL,孵育60 min降至3.33×102 PFU/mL,在90℃孵育30 min后,噬菌体活性完全丧失(图7)。
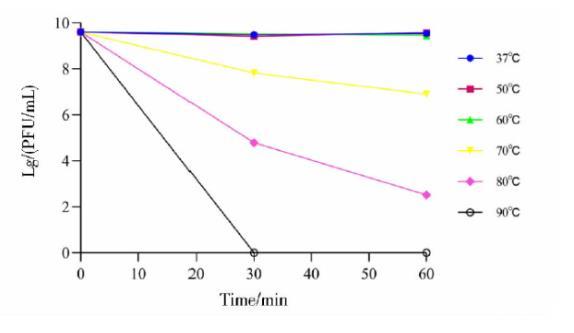
图7噬菌体PJNB028热稳定性
2.7噬菌体PJNB028的pH稳定性测定
如图8所示,噬菌体PJNB028在pH为3~11的缓冲液中处理1 h效价无明显变化,在pH>2和pH<12时完全失活。
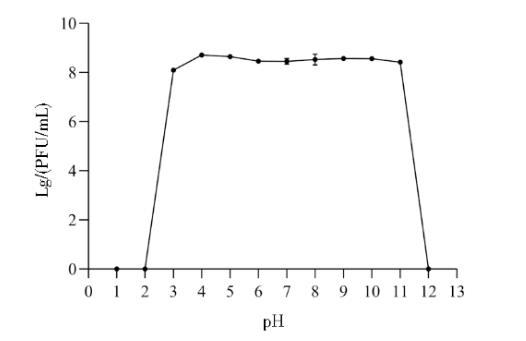
图8噬菌体PJNB028的pH稳定性
2.8噬菌体PJNB028紫外线敏感性测定
如图9所示,噬菌体PJNB028在紫外灯照射下效价逐渐降低,效价从1.5×109 PFU/mL降低为1.36×104 PFU/mL,表明噬菌体PJNB028对紫外线较敏感。
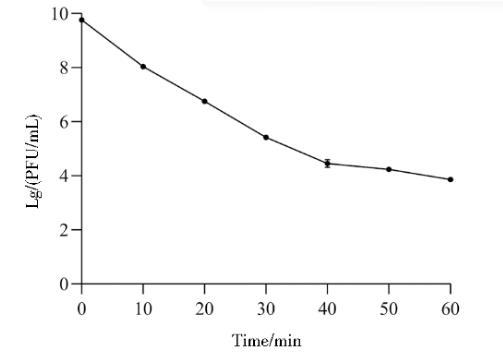
图9噬菌体PJNB028紫外线照射稳定性
2.9噬菌体PJNB028体外抑菌效果测定
如图10所示,当MOI为0.001~10时,不同浓度噬菌体均能对细菌生长起到一定抑制作用,抑制时间在1~8 h,当MOI为1时对细菌生长的抑制效果最好。
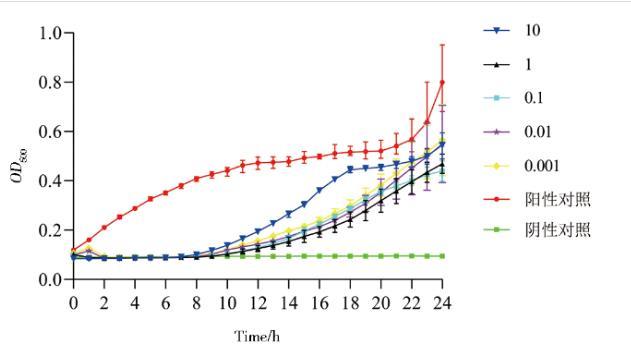
图10噬菌体PJNB028的体外抑菌效果测定
2.10噬菌体PJNB028裂解谱测定
如表2所示,噬菌体PJNB028对5株莫哈韦芽孢杆菌、5株枯草芽孢杆菌具有裂解性,对其余8株枯草芽孢杆菌、1株蜡样芽孢杆菌和1株地衣芽孢杆菌无裂解性。
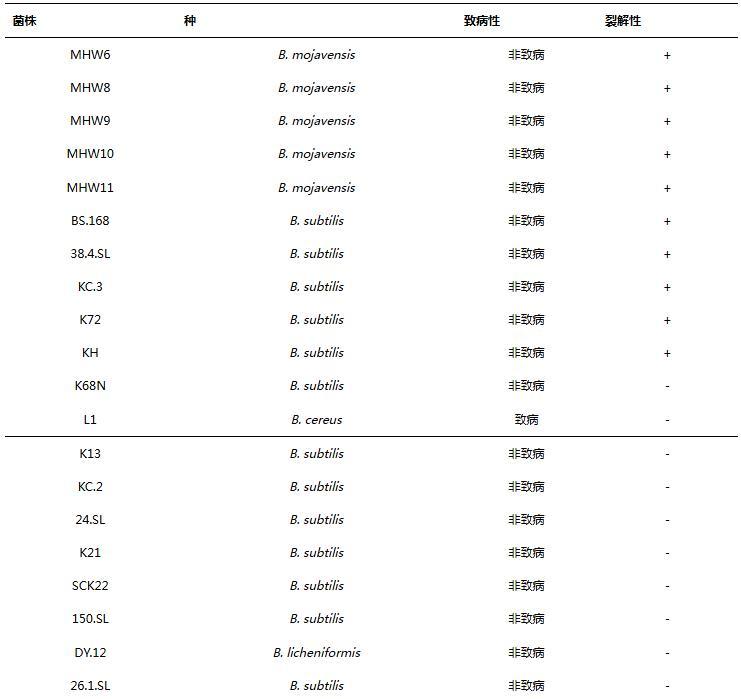
表2噬菌体PJNB028宿主谱
注:+表示裂解,-表示不裂解
2.11抗性菌株筛选
使用宿主菌(图11A)筛选到3株不能裂解的抗性菌株(图11B),如图11C所示,对数期的宿主菌和抗性菌与噬菌体共培养5 h后,宿主菌变澄清,抗性菌与噬菌体双层板未见到噬菌斑(图11D)。在显微镜下观察,被噬菌体裂解的宿主菌体明显稀少(图11F),而抗性菌株菌体未发现明显变化。
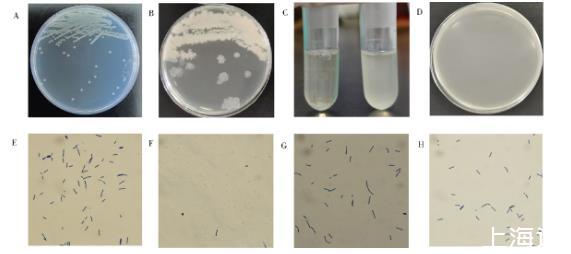
图11宿主菌与抗噬菌体菌株的比较
A:宿主菌株菌落形态B:抗噬菌体突变菌株菌落形态C:加入噬菌体共培养5 h后菌液对比,左为宿主菌,右为抗性菌株D:抗性菌株与噬菌体双层平板结果E:宿主菌对数期菌液形态F:对数期宿主菌加入噬菌体共培养5 h后菌液形态G:抗性菌株对数期菌液形态H:对数期抗性菌株加入噬菌体共培养5 h后菌落形态
2.12菌体生长动态
经48 h发酵培养监测显示(图12),野生型菌株T0与突变株T1和T3的OD600值呈现显著增长态势,而T2的生长速率较缓。
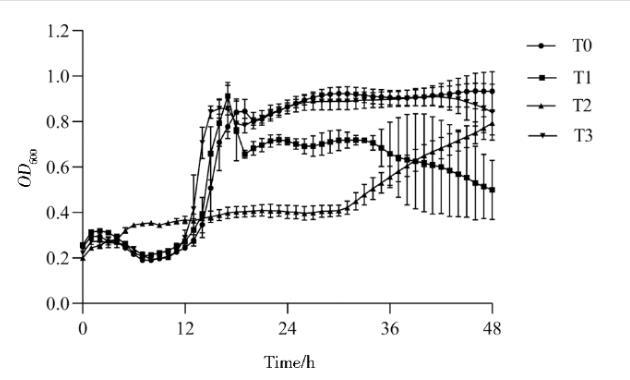
图12发酵菌株生长曲线
2.13菌液效价随时间的变化
如图13所示,抗性突变株T1和T3的效价表现更优:发酵24 h时,T3效价为(8.9±0.1)Lg(CFU/mL),显著高于野生型T0;而T2在发酵24 h和36 h时的效价均显著低于T0。不同突变菌株的生长特征不同,有的优于野生型,有的比野生型菌株弱。
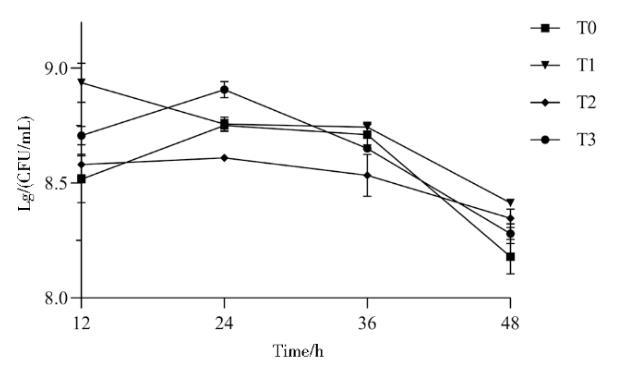
图13发酵菌株效价变化
2.14淀粉酶活性分析
在发酵过程中,T0和T3的酶活在24 h达到峰值,随后显著下降,如图14所示。值得注意的是,菌株T2呈现出独特的持续增长趋势,其酶活在48 h内从12 h的(168±1)μg/(min·mL)呈线性上升至(452±1)μg/(min·mL)。此外,菌株T3在24 h时的酶活水平显著高于其他突变株。不同时间段菌株产生的淀粉酶活性不同,菌株的代谢能力不同。
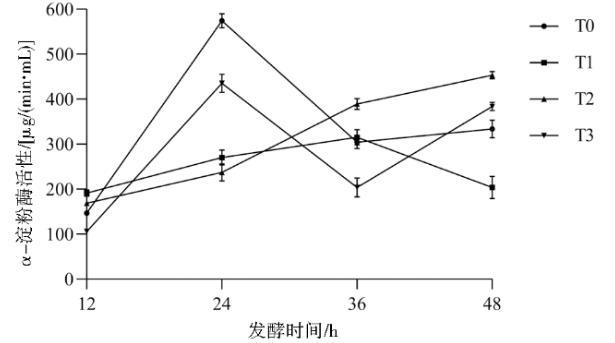
图14α-淀粉酶活性动态变化
3讨论
本研究从高温堆肥中分离纯化出一株枯草芽孢杆菌噬菌体PJNB028,可以在枯草芽孢杆菌上形成2 mm完全透亮的噬菌斑,噬菌斑周围有晕环,随着孵育时间增加,晕环也逐渐增大,推测该噬菌体可能分泌噬菌体衍射酶。
根据系统发育分析PJNB028噬菌体属于Caudoviricetes纲,与2S-4、PK-3最近,与16株噬菌体同源性较高,表明这些噬菌体都是潜在的已经分离到的污染源,PJNB028具有代表意义。该噬菌体的最佳MOI为1,增殖潜伏期为10 min,爆发期为70 min,爆发量为291 PFU/cell,基因组大小为162 308 bp,含有237个ORF。通过全基因组分析和生物学研究发现其具有良好的生物学特性,属于枯草芽孢杆菌噬菌体的优势噬菌体。
噬菌体PJNB028基因组中有2个tRNA基因与Arg(细菌精氨酸)稀有密码子对应,携带tRNA的噬菌体能够减少对宿主菌tRNA的依赖,一定程度上增强裂解能力从而扩大裂解谱范围。噬菌体受体结合蛋白(RBPs)是决定噬菌体裂解谱的一个关键因素,噬菌体可通过ORF138尾钉蛋白(tail spike proteins,TSPs)上的RBPs吸附在宿主菌表面进行裂解,有研究表明RBP可作为荧光纳米探针识别宿主细菌。ORF208毒素-抗毒素(toxin-antitoxin,TA)系统是原核生物重要免疫系统之一,在细菌抵抗噬菌体防御机制中占有主导作用。噬菌体在与TA系统协同进化过程中进化出对TA系统的防御,并且噬菌体可利用细菌的毒素-抗毒素系统控制裂解或溶原的状态。
噬菌体PJNB028较耐高温,在80℃处理1 h效价明显降低,说明通过高温可减少噬菌体的污染。Krasowska等分离的枯草芽孢杆菌噬菌体耐高温,在121℃条件下孵育1 min仍保持感染活性,一般认为Myoviridae和Siphoviridae家族成员对较大温度波动具有抵抗力。在紫外线照射下,噬菌体的效价降低较缓慢,表明该噬菌体对野外紫外线较耐受,而赵欣卓等分离的枯草芽孢杆菌噬菌体Bac-S在紫外线下21 min完全失活。噬菌体PJNB028在pH为3~11时效价稳定,耐酸不耐碱,可以热碱液消灭。与一些对枯草芽孢杆菌的研究结果不同,Zhang等分离的枯草芽孢杆菌噬菌体phi18耐碱不耐酸(pH 4~12)。
通过自然选择或基因工程可以制备抗噬菌体突变体,刘秀侠等为防止噬菌体P260污染,利用自发突变的方法筛选出一株优良的枯草芽孢杆菌突变菌。张建璀通过自然选育和紫外诱变育种选育的方法,筛选出噬菌体TXp1的抗噬菌体菌株。本研究使用自然突变的方法共培养筛选得到抗性菌株3株,与野生菌株进行对比试验发现部分菌株在发酵过程中表现出更高的α-淀粉酶活性及菌株效价,为工业发酵菌株改良提供了新策略。
4结论
目前,约60%商用酶是由芽孢杆菌产生的,但在工业发酵中噬菌体感染不可避免,同时,部分枯草芽孢杆菌也是引起食品腐败、感染的有害细菌,而枯草芽孢杆菌噬菌体也可能会在预防食品腐败和其他医疗方面有用途。本文分离的枯草芽孢杆菌裂解性噬菌体PJNB028属于Caudoviricetes纲,具有较宽裂解谱,通过噬菌体与宿主菌共培养筛选到3株抗菌菌株;配合高温蒸汽、热碱液可以彻底消毒。从有益芽孢杆菌和致病芽孢杆菌防治两方面看,进一步建立枯草芽孢杆菌及其噬菌体资源库、共进化培养及基因编辑平台具有巨大的需求和应用潜力。
相关新闻推荐
1、一株具有高渗透压耐受性的嗜果糖植物乳杆菌的分离与特性研究(一)
3、分枝杆菌最低抑菌浓度测定、PASTA蛋白功能表征及膜蛋白组学分析