CRISPR-Cas系统是原核生物中的一种适应性免疫系统,用于抵御外来核酸的入侵,如噬菌体和质粒。CRISPR-Cas系统分为两大类,其中III型系统因其独特的机制和能够同时切割入侵者的RNA和DNA而受到关注。III型系统进一步分为多个亚型,其中III-E型系统与其他亚型存在显著差异,其效应蛋白gRAMP与一个类似半胱氨酸蛋白酶的TPR-CHAT结构域结合形成Craspase复合物,但其具体的免疫机制尚不清楚。研究人员利用冷冻电镜技术解析了gRAMP-crRNA复合物以及Craspase复合物与靶标RNA(CTR和NTR)结合时的结构,分辨率达到原子级别。研究人员还通过体外RNA切割实验、质谱分析、蛋白质相互作用实验以及细菌生长实验(Bioscreen C)等多种方法,研究了Craspase复合物的功能和作用机制。Bioscreen C全自动微生物生长曲线分析仪被用于细菌生长实验,并提供了实时、高通量的细菌生长监测,还帮助研究人员评估了III-E型CRISPR-Cas系统的抗病毒效果,为理解其免疫机制提供了重要的实验数据支持。研究发现Craspase复合物由gRAMP和TPR-CHAT组成,gRAMP包含多个Cas7样结构域和一个Cas11样结构域,而TPR-CHAT包含一个TPR结构域和一个CHAT结构域。Craspase复合物的结构表明,TPR-CHAT与gRAMP的Cas7.2结构域相互作用,锁定crRNA的5'标签区域。实验表明,当Craspase复合物结合与crRNA互补的靶标RNA(CTR)时,TPR-CHAT的蛋白酶活性被激活,能够特异性地切割辅助蛋白Csx30。这种切割事件会触发一种称为“自杀性感染”的抗病毒策略,即细胞通过自我牺牲来阻止病毒的传播。
Bioscreen全自动生长曲线分析仪的应用
Bioscreen全自动生长曲线分析仪被用于细菌生长实验。使用非诱导的过夜培养的细菌(大肠杆菌,携带编码gRAMP-RNA、CHAT和Csx30-Csx31-SbRpoE的质粒)或阴性对照(携带相应空载体的大肠杆菌)。将这些细菌培养物稀释至最终OD600 nm为0.4的LB培养基中,培养基中补充了氨苄青霉素、卡那霉素和氯霉素。每个180µL的培养物被转移到一个100孔板中,每个孔中加入18µL的噬菌体裂解液(或18µL的SM缓冲液作为未感染对照),最终噬菌体λ的感染复数(MOI)为4和0.1。向每个孔中加入2µL的10 mM IPTG,使最终IPTG浓度达到0.1 mM。感染实验以三份重复进行,每份重复来自单独的过夜培养物。将蜂窝平板置于Bioscreen C中,37°C下振荡培养。每15分钟测量一次OD600 nm的吸光度,以监测细菌的生长情况。Bioscreen C仪器能够在同一实验中同时处理多个样本,提高了实验效率,使得研究人员能够快速获得大量数据,从而更准确地评估不同条件下的细菌生长和抗病毒效果。
实验结果
III-E型CRISPR-Cas系统的Craspase复合物通过识别靶标RNA的非互补性来激活其蛋白酶活性。Csx30的特异性切割是触发自杀性感染的关键步骤,这一过程需要Csx31和SbRpoE的参与。该系统通过自杀性感染机制来抵御病毒侵染,为理解原核生物的免疫机制提供了新的视角,并为开发新型抗病毒工具提供了可能的途径。通过在大肠杆菌中表达III-E型CRISPR-Cas系统,并利用噬菌体λ进行感染实验,证明了该系统能够有效抵御噬菌体的侵染。Csx30、Csx31和SbRpoE在抗病毒活性中起着关键作用。缺失这些组分或使用非靶向的CRISPR阵列会显著降低或消除对噬菌体的保护效果。
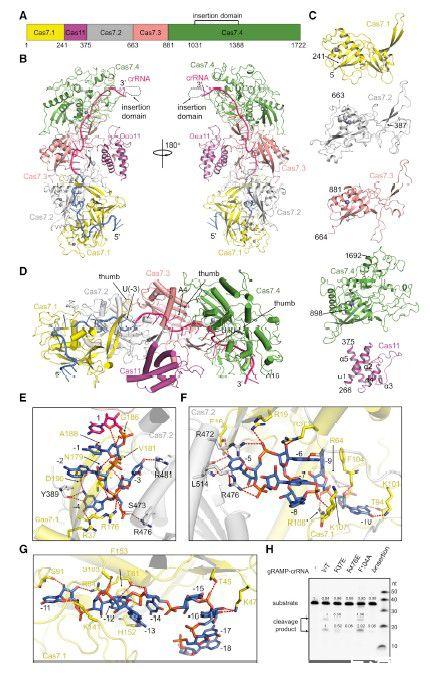
图1、gRAMPcrRNA的冷冻电镜结构。(A)Candidatus Scalindua brodae中gRAMP的结构域结构。Cas7.1–7.4和Cas11结构域分别以洋红色、黄色、灰色、粉红色和绿色着色。(B)gRAMP-crRNA的整体结构。gRAMP的域的颜色如(A)所示。crRNA的颜色为亮粉色和海洋色,分别用于间隔区和重复区。标记了Cas7.4中的未建模插入域。(C)gRAMP的单独结构域的结构。二级结构标记为Cas11结构域。(D)crRNA结合的特写视图。标记了Cas7.1-7.3结构域的拇指和crRNA中的扭结核苷酸。(E-G)gRAMP的Cas7.1/7.2结构域与crRNA的核苷酸(1)–(4)和crRNA(F)的(5)–(10)或(11)–(18)之间的相互作用。红色虚线表示极相互作用。(H)gRAMP-crRNA及其突变体的靶RNA切割。DInsertion表示插入结构域(残基1,031-1,388)缺失并被GSG(甘氨酸-丝氨酸甘氨酸)接头取代的gRAMP-crRNA。
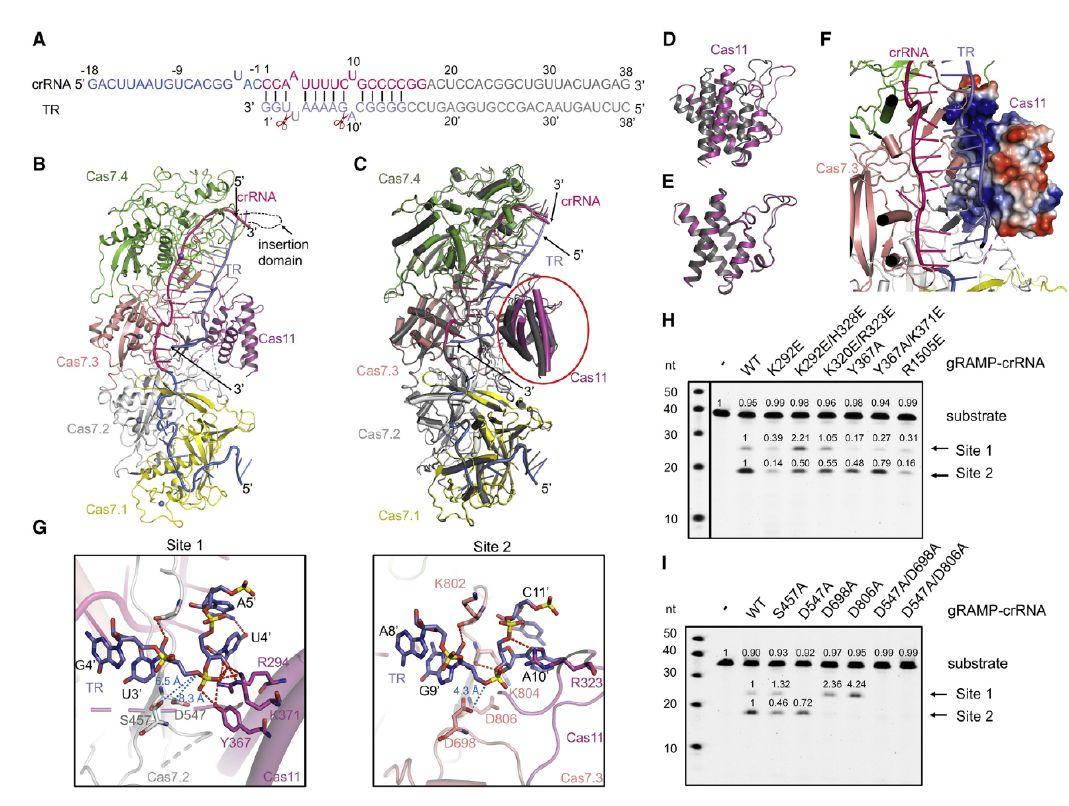
图2、gRAMP-crRNA-TR和RNA切割机制的结构。(A)crRNA-TR双链体的示意图。实验中使用的crRNA和TR核苷酸用线条表示。TR是石板色的。crRNA和TR的建模核苷酸如结构所示。不可见的核苷酸呈灰色。剪刀表示卵裂部位。(B)gRAMP-crRNA-TR的整体结构。TR的颜色与(A)相同。(C)gRAMP-crRNA(深灰色)和gRAMP-crRNA-TR(B)的结构叠加。Cas11结构域的区域以圆圈突出显示。(D)(C)结构对齐内Cas11结构域区域的特写图。(E)gRAMP-crRNA和gRAMP-crRNA-TR中独立Cas11结构域的结构对齐。(F)gRAMP-crRNA-TR结构中Cas11区域的特写视图,其中Cas11结构域在静电模型中是彩色的。(G)两个切割位点gRAMP和TR之间的详细相互作用。红色虚线表示极相互作用。(H)gRAMP-crRNA及其突变体的靶RNA切割损害间隔区TR结合。标记了代表位点1和2切割的产物条带。(I)gRAMP-crRNA的靶RNA切割及其在两个切割活性位点发生突变的突变体。
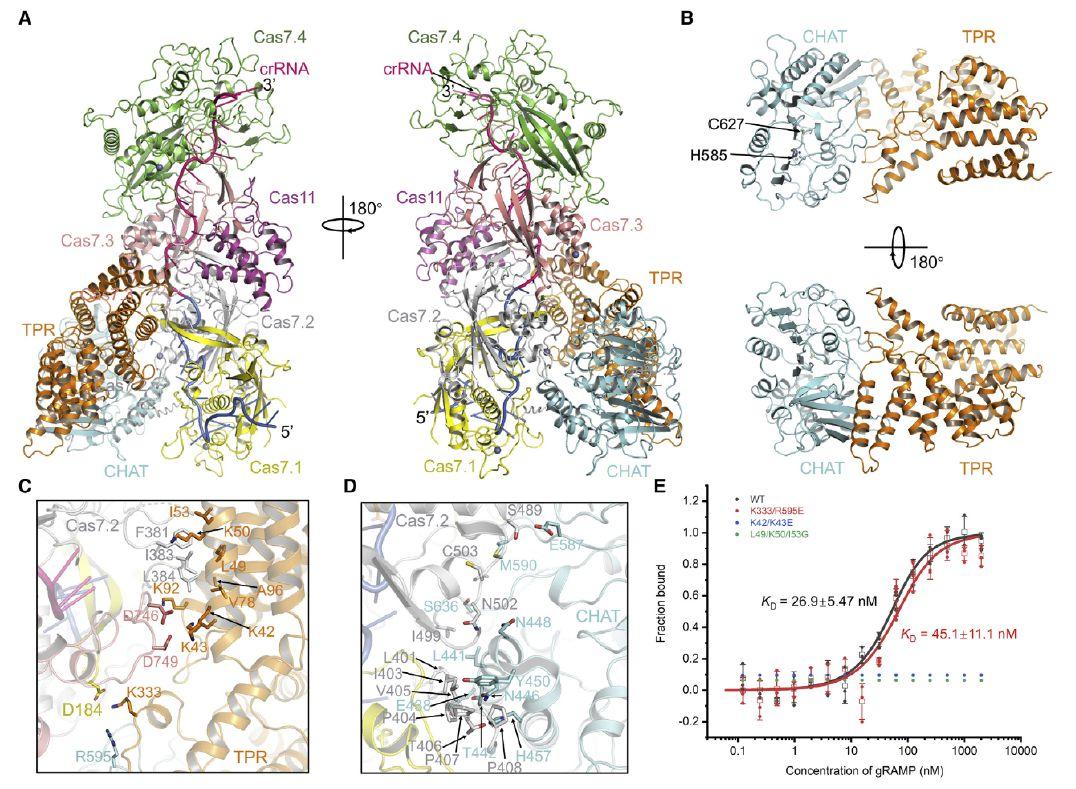
图3、Craspase的冷冻电镜结构。(A)Craspase的整体结构。gRAMP-crRNA的颜色如图1B所示。TPR和CHAT结构域的TPR-CHAT分别为橙色和青色。(B)Craspase中TPR-CHAT的整体结构,颜色如(A)。两个活性位点残基以棒状显示。(C和D)gRAMP与TPR-CHAT的TPR域(C)和CHAT域(D)之间的详细交互。(E)gRAMP-crRNA与TPR-CHAT及其突变体结合的MST测定。显示了来自三个独立实验的单个值。
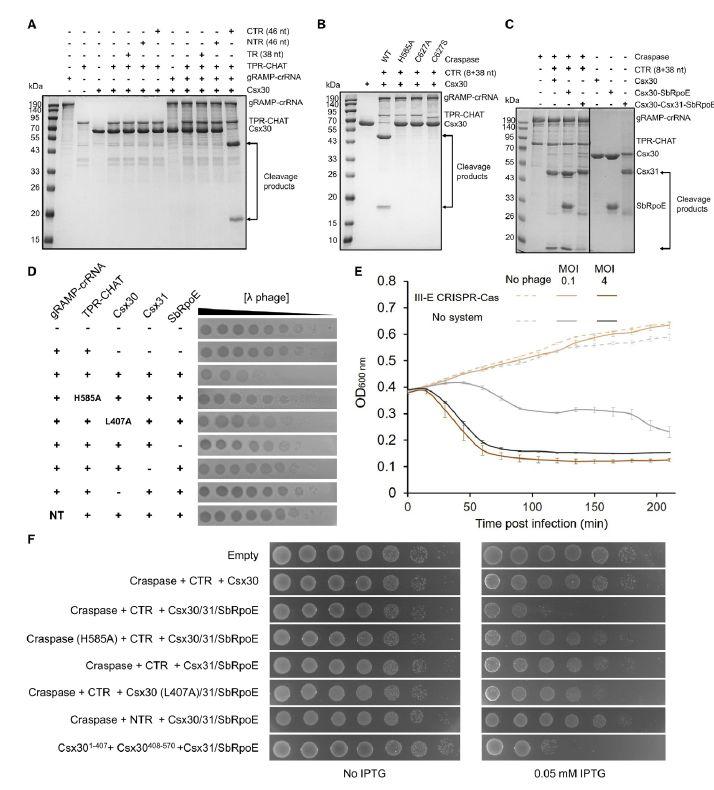
图4、III-E型CRISPR-Cas系统的免疫机制。A)Craspase复合物中TPR-CHAT切割的靶标RNA依赖性Csx30。TR/NTR/CTR RNA分子、TPR-CHAT、gRAMP-crRNA和Csx30以0.5、0.3、0.3和1.5 mM的浓度添加。(B和C)靶标RNA依赖性Csx30与Craspase或其突变体在TPR-CHAT的活性位点发生突变。(D)III-E型CRISPR-Cas纵子可防止噬菌体。显示了在表达来自Candidatus Scalinduabrodae的III-E CRISPR-Cas纵子型基因的大肠杆菌BL21(DE3)细胞上接种l噬菌体的效率。该实验已独立重复3次,并显示了具有代表性的结果。(E)表达III-E CRISPR-CAS型纵子的五个基因的大肠杆菌细胞或对照大肠杆菌细胞(无系统)的大肠杆菌细胞的液体培养物的生长曲线(Bioscreen)。细胞在37C下用l噬菌体感染。细菌在时间0以4或0.1的MOI感染。(F)III-E型CRISPR-Cas系统的激活是有毒的。将编码指定质粒的细胞以10倍连续稀释接种在溶原肉汤(LB)-琼脂上,在没有诱导或诱导表达的条件下接种。
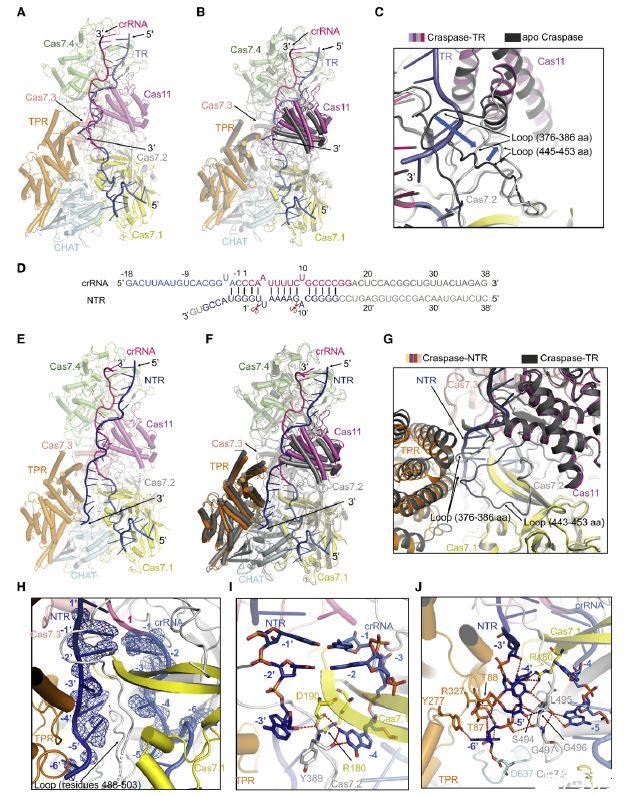
图5、Craspase-TR和Craspase-NTR的冷冻电镜结构(A)与TR(彩色板岩)结合的Craspase(如图3A所示)的整体结构。(B)Craspase(深灰色)和Craspase-TR(颜色如A所示)的结构叠加。(C)(B)中结构叠加中与Cas11结构域相邻的两个环的特写图。Craspase和Craspase-TR的颜色如(B)。(D)Craspase-NTR中crRNA-NTR双链体的示意图。实验中使用的crRNA和NTR核苷酸用线条表示。NTR为深蓝色。不可见的核苷酸呈灰色。(E)与NTR(深蓝色)结合的Craspase的整体结构。
总结
本研究主要是关于一种新型的CRISPR-Cas系统——III-E型CRISPR-Cas系统的免疫机制研究。研究人员通过结构生物学和生物化学方法,揭示了该系统中Craspase复合物的结构特征以及其在抗病毒防御中的作用机制。在III-E型CRISPR-Cas系统中,Cas效应蛋白(gRAMP)与一个TPR-CHAT(tetratricopeptide repeat-Caspase HetF associated with TPRS)结合形成Craspase(CRISPR引导的半胱氨酸蛋白酶)。然而gRAMP的结构特征以及该系统的免疫机制尚不清楚。在此,研究人员解析了gRAMP-crRNA以及gRAMP:crRNA:靶标RNA的结构,同时解析了Craspase以及Craspase与靶标RNA(CTR或NTR)复合物的结构。重要的是,NTR和CTR的3'反标签区域结合在Craspase的两个不同通道中,且CTR的非互补3'反标签能够诱导TPR-CHAT发生显著的构象变化,从而别构激活其蛋白酶活性,进而切割辅助蛋白Csx30。这种切割会触发III-E型系统的自杀性感染作为抗病毒策略。本研究不仅揭示了Craspase复合物的结构特征,还阐明了其在III-E型CRISPR-Cas系统中的作用机制,特别是靶标RNA如何通过非互补性激活蛋白酶活性。研究揭示了III-E型CRISPR-Cas系统通过自杀性感染来抵御病毒侵染的独特机制,为理解原核生物的免疫机制提供了新的视角。这些发现为开发新型抗病毒工具和基因编辑技术提供了理论基础,特别是在设计能够特异性切割RNA的工具方面具有重要意义。Bioscreen C仪器能够实时记录细菌在不同时间点的OD600 nm吸光度,从而反映细菌的生长曲线。实现了实时监测细菌生长,评估抗病毒效果,通过比较携带CRISPR-Cas系统的细菌与携带空载体的对照细菌在噬菌体感染后的生长差异,这使得研究人员能够动态观察细菌在感染噬菌体后的生长变化,以及III-E型CRISPR-Cas系统对噬菌体感染的防御效果。
相关新闻推荐
1、畜禽屠宰场环境和畜禽肉中单增李斯特菌污染情况、生长曲线及消毒剂最小抑菌浓度分析(三)