潜在致病菌的生长。
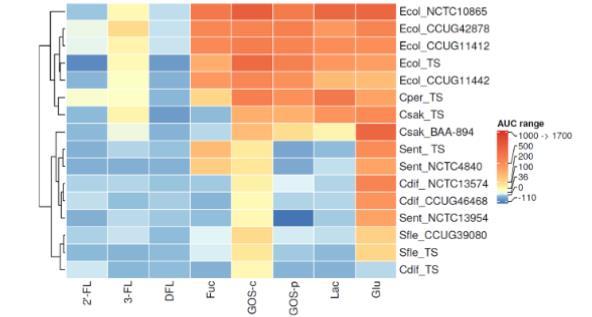
图4. 碳源对潜在致病菌生长的影响。通过生长曲线下面积(AUC)评估添加1% 2'-岩藻糖基乳糖(2'-FL)、3-岩藻糖基乳糖(3-FL)、二岩藻糖基乳糖(DFL)、岩藻糖(Fuc)、商业低聚半乳糖(GOS-c)、富集低聚糖的GOS(GOS-p)、乳糖(Lac)和葡萄糖(Glu)后潜在致病菌的生长情况。AUC值通过减去未添加碳水化合物的生长培养基中细菌的AUC值进行标准化。生长数据采用色标标注:深红色代表强烈生长(AUC值范围:1000-1700;500-999;200-499;100-199;36-99;0-35),深蓝色代表生长低于培养基基准(AUC值范围:-110至-1)。细菌菌株缩写见表2。
潜在致病菌的碳水化合物利用结果以热图形式显示在图4中。与上文相同,每种细菌的每种碳源的AUC值减去了相应未添加碳水化合物的培养基的AUC值,并根据生长模式的相似性对细菌菌株进行聚类。
16种潜在致病菌形成了两个主要簇。聚类受到所有研究的大肠杆菌菌株和产气荚膜梭菌模式菌株(Cper_TS)对岩藻糖的利用以及在3-FL上的轻微生长,以及阪崎克罗诺杆菌模式菌株(Csak_TS)在3-FL上的轻微生长的影响。此外,上述所有物种和阪崎克罗诺杆菌BAA-94(Csak_BAA-894,一种人类临床分离株)在GOS-c、GOS-p、乳糖和葡萄糖上生长良好。
2'-FL和DFL不支持任何潜在致病菌的生长。我们观察到大肠杆菌、产气荚膜梭菌和阪崎克罗诺杆菌在3-FL上有极弱或弱生长,这在生长实验开始时观察到,并缓慢下降,在24小时时与培养基对照相比几乎无差异。这可能是受到3-FL产品中少量残留乳糖和半乳糖(见表1)的影响。
表1:碳源的结构、成分和供应商| 碳水化合物 | 类型 | 缩写 | 纯度 | 供应商 |
|---|---|---|---|---|
| 2'-岩藻糖基乳糖, Fuc a1-2 Galβ1-4 Glu | 三糖 | 2'-FL | 2'-FL 99% | 杜邦营养与生物科学公司 |
| 3-岩藻糖基乳糖, Galβ1-4 Glu a1-3 Fuc | 三糖 | 3-FL | 3-FL 96%, 乳糖 2.5%, 半乳糖 0.3% | 杜邦营养与生物科学公司 |
| 二岩藻糖基乳糖, Fuc a1-2 Galβ1-4 Glu a1-3 Fuc | 四糖 | DFL | DFL 97%, 3-FL 0.1%, 2'-FL 2.1% | 杜邦营养与生物科学公司 |
| 岩藻糖 | 单糖 | Fuc | 岩藻糖 99.5% | 杜邦营养与生物科学公司 |
| Bimuno GOS | 多糖 | GOS-c | GOS 81%, 葡萄糖 5.4%, 乳糖 12.3% | Clasado Biosciences |
| 乳糖 | 二糖 | Lac | 用于微生物学 | Merck |
| 葡萄糖 | 单糖 | Glu | ACS级 | J.T. Baker |
注:GOS=低聚半乳糖;Gal=半乳糖
几种肠杆菌科能很好地利用单糖和二糖,这表明存在与HMOs利用者(尤其是两歧双歧杆菌和拟杆菌属)交叉喂养的可能性,这些菌株在细胞外降解HMO结构并释放较小的单糖、二糖和寡糖。或者,已知产气荚膜梭菌含有1,3-1,4-α-L-岩藻糖苷酶,能够从粘蛋白中释放岩藻糖,表明其具有切割岩藻糖的能力,也可能作用于3-FL。相反,Yu等人未观察到大肠杆菌、产气荚膜梭菌或肠杆菌属在48小时后3-FL与培养基对照之间的任何差异。
所有大肠杆菌菌株和产气荚膜梭菌模式菌株(Cper_TS)利用了岩藻糖,而两种阪崎克罗诺杆菌菌株则没有,这与先前的研究结果一致。阪崎克罗诺杆菌是配方奶粉中常见的病原体。观察到大肠杆菌菌株对岩藻糖利用的变异性;模式菌株(Ecol_TS,来自尿液)和菌株NCTC10865(Ecol_NCTC10865,来自腹泻婴儿的粪便)在岩藻糖上生长良好,但不如在葡萄糖上好。菌株CCUG11412(Ecol_CCUG11412,来自婴儿肠胃炎)和CCUG42878(Ecol_C-CUG42878,来自婴儿肠胃炎)对葡萄糖和岩藻糖的利用程度相同,而菌株CCUG11442(Ecol_C-CUG11442,来自婴儿腹泻)在岩藻糖上的生长优于葡萄糖。因此,大肠杆菌菌株之间在利用岩藻糖的能力上存在差异。岩藻糖利用已被认为对致病性大肠杆菌EHEC菌株的定植至关重要;因此,两歧双歧杆菌和拟杆菌属对HMOs和其他聚糖的利用(它们在细胞外切割岩藻糖)可能有助于EHEC的毒力。
此外,在本研究中,沙门氏菌菌株对岩藻糖的消耗情况各不相同。三种沙门氏菌菌株中有两种在岩藻糖上生长:鼠伤寒沙门氏菌肠道亚种模式菌株(Sent_TS,来源未知)和波那沙门氏菌肠道亚种菌株NCTC4840(Sent_NCTC4840,来自婴儿肠炎),而鼠伤寒沙门氏菌肠道亚种菌株NCTC13954(Sent_NCTC13954,来自人类粪便)则没有。先前的研究表明,所有已知的沙门氏菌肠道菌株都含有岩藻糖利用操纵子。我们发现菌株NTCT13954(血清型鼠伤寒)未在岩藻糖上生长,这表明岩藻糖操纵子的表达存在变异性,至少在实验条件下是如此。
所有大肠杆菌菌株、产气荚膜梭菌模式菌株(Cper_TS)和两种阪崎克罗诺杆菌菌株在GOS-c和GOS-p上都生长良好。这些结果与早期显示这些细菌在GOS和乳果糖上生长的一项研究一致,以及其他显示大肠杆菌和产气荚膜梭菌在GOS和乳糖上生长的研究。然而,另一项研究未发现大肠杆菌在GOS上生长。总体而言,在我们的研究中,大肠杆菌在商业和纯化GOS上都生长良好,并且根据菌株的不同,生长优于或等于在葡萄糖上观察到的生长。所有在葡萄糖上生长并在GOS-c上微弱生长的艰难梭菌、福氏志贺氏菌和沙门氏菌菌株均未在GOS-p上生长,GOS-p含有很少的单糖和二糖,表明它们无法利用纯GOS产品。此外,无法在GOS-p上生长通常伴随着无法在乳糖上生长。所有研究的大肠杆菌菌株都在乳糖上生长。早期的研究表明,非致病性大肠杆菌菌株不利用乳糖,而模式菌株已被证明可以利用乳糖。
福氏志贺氏菌的结果与先前一项研究的结果相似,该研究发现痢疾志贺氏菌仅在葡萄糖和GOS上生长。肠杆菌科对不同GOS产品利用的改变也已被观察到。先前很少有研究评估潜在致病菌在GOS等益生元上的生长。不同GOS产品的组成各不相同,这也可能导致细菌菌株在利用GOS方面的依赖性差异。
在最近的一个单一培养模型中,乳糖和GOS都增加了产气荚膜梭菌的生长。艰难梭菌模式菌株(Cdif_TS)的总体生长较弱,使得与其他菌株进行比较变得不可能。
岩藻糖的利用将这群潜在致病菌分为两个簇。在2'-FL和DFL上未发现生长,而一些细菌可能能够利用3-FL,尽管效率不高。利用岩藻糖基化HMO需要特定的酶,特别是1,2-a-L-岩藻糖苷酶,这可以在一些特定的婴儿共生菌中找到。HMOs作为选择性营养素,可以保护婴儿免受病原体侵害。从GOS中去除大部分单糖和二糖阻止了沙门氏菌属和志贺氏菌属的生长。
所有细菌的聚类和结论。
为了可视化所有研究的57种细菌的聚类,对所有处理的相关矩阵进行了主成分分析(图5)。提取了前四个主成分,它们解释了数据中95%的方差。基于提取的主成分将菌株聚类为六个簇。
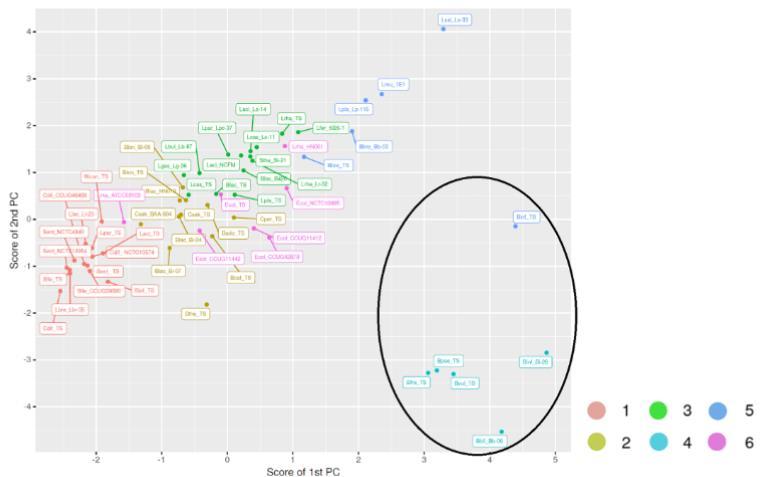
图5. 基于测试碳水化合物生长的细菌聚类分析:2'-岩藻糖基乳糖、3-岩藻糖基乳糖、二岩藻糖基乳糖、岩藻糖、乳糖、葡萄糖,以及商业级与纯化形式的低聚半乳糖(GOS)。
然而,当所有各种细菌物种合并时,HMO的利用是决定簇的主要因素。簇4包含在岩藻糖基化HMOs上明显生长的细菌:婴儿双歧杆菌Bi-26(Binf_Bi-26)、两歧双歧杆菌Bb-06(Bbif_Bb-06)、假小链双歧杆菌模式菌株(Bpse_TS)、脆弱拟杆菌模式菌株(Bfra_TS)和普通拟杆菌模式菌株(Bvul_TS)。簇2、3和5都包含未在HMOs和岩藻糖上生长但在所有其他研究的碳源上生长的细菌。这些簇之间的差异与簇5中在葡萄糖、乳糖和两种GOS产品上的强生长有关;簇3中在纯化GOS上的生长相比商业GOS减少;以及簇2中在葡萄糖、乳糖和GOS上的总体生长减少。婴儿双歧杆菌模式菌株(Binf_TS)也被包括在簇5中,可能是由于其在GOS上的强生长。然而,该物种在图表中被分组到更靠近其他HMO利用者,而不是那些无法利用HMOs的物种。簇6包含利用岩藻糖的细菌,簇1包括要么总体生长低要么主要在葡萄糖上生长的细菌。
一些碳水化合物比较研究使用了贫瘠培养基,例如ZMB1,以避免复杂的培养基成分(包括酵母或牛肉提取物)可能通过在没有碳源的情况下为背景生长提供营养而影响结果。因为我们评估了具有不同营养需求的异质细菌群,我们希望细菌在各自的最佳培养基中生长,这允许在组内和组间进行比较。我们的主要目的是检查细菌生长的差异,我们研究了三种HMOs、其他常用益生元以及寡糖组分对大量双歧杆菌、乳杆菌科和潜在致病菌的影响。评估纯培养物生长的方法既可靠又可重复,但也存在差异。Yu等人报告了48小时细菌生长后600 nm处光密度的变化,Hoeflinger等人报告了24-96小时细菌生长期间600 nm处最大光密度的变化。我们选择使用AUC来评估每种细菌菌株的生长,因为该参数还考虑了延迟时间、总体生长强度和生长速率。将数据呈现为AUC值也使得更容易在各种碳源之间进行比较。对纯细菌培养物在各种底物存在下生长的深入了解使得能够估计在更复杂条件下碳水化合物的利用和交叉喂养。将来,可以利用有限数量的细菌物种/菌株组合使用碳水化合物混合物来研究交叉喂养动态。
目前的工作强调了评估益生菌和病原菌菌株特异性差异的重要性。由于HMOs越来越多地被添加到婴儿配方奶粉中,将HMOs的益生元效果与其他益生元(如GOS)的效果进行比较变得更加重要。
HMOs的利用增加了母乳喂养婴儿中双歧杆菌的丰度。运输和降解岩藻糖基化HMO结构的能力对于这一过程至关重要。目前的工作支持这一模型,因为只有选定的双歧杆菌和拟杆菌利用了岩藻糖基化HMOs,而其他研究的细菌(例如病原菌)则没有。还观察到潜在致病菌在岩藻糖和GOS利用方面的差异。
这些结果显示了与HMOs相关的选择性以及各种益生菌和病原菌之间益生元消耗的差异。HMOs的选择性可能有利于婴儿肠道微生物群的进化,通过促进有益微生物的生长,同时不支持有害病原体的生长,这与其他选择性较差的益生元形成对比。
相关新闻推荐
1、减毒重组沙门氏菌递送鼠疫耶尔森菌多抗原诱导完全保护性免疫