研究简介
本研究旨在探讨环丙烷脂肪酸(CFA)修饰在鼠伤寒沙门氏菌(Salmonella enterica serovar Typhimurium)应激抵抗和毒力中的关键作用。CFA是一种常见的膜脂质修饰方式,广泛存在于真细菌、植物和原生动物中,由cfa基因编码的CFA合酶催化形成。先前研究表明,CFA与细菌对氧化应激、有机溶剂和酸应激的抵抗相关,但其在沙门氏菌致病机制中的具体贡献尚不明确。多种真细菌、植物和原生生物能够通过环丙烷化修饰膜脂,这种修饰据报道可以调节膜的通透性和流动性。环丙烷化膜脂的能力已被发现与多种应激耐受性相关:在结核分枝杆菌中与氧化应激耐受性相关,在大肠杆菌中与有机溶剂应激耐受性相关,在大肠杆菌和沙门氏菌中与酸性应激耐受性相关。
在细菌中,编码环丙烷脂肪酸(CFA)合酶的cfa基因在生长的稳定期(stationary phase)会被诱导表达。本论文研究构建了鼠伤寒沙门氏菌14028s(S.Typhimurium)cfa突变株,并研究了CFA修饰的脂质在应激耐受性及小鼠体内毒力中的作用。我们对野生型和cfa突变株的CFA含量进行了定量分析,结果显示cfa突变株的CFA水平显著低于野生型,表明CFA合酶是沙门氏菌中环丙烷化脂质的主要催化酶。cfa突变株在极酸性pH条件下、质子载体CCCP存在下以及过氧化氢暴露时,对应激更加敏感。此外,cfa突变株在小鼠巨噬细胞中存活能力下降,而添加NADPH吞噬细胞氧化酶抑制剂二苯基碘鎓(DPI)可部分恢复其存活能力。缺失cfa的S.Typhimurium在小鼠体内毒力也显著下降。这些结果表,CFA修饰的脂质在沙门氏菌毒力中发挥重要作用。
Bioscreen全自动生长曲线分析仪的应用
将野生型鼠伤寒沙门氏菌(ATCC 14028s)及其同源cfa突变体培养至稳定期,用LB培养基稀释至OD600为0.002。在实验组中加入1 mM过氧化氢(H₂O₂),对照组不加应激剂,以模拟氧化应激条件。使用Bioscreen C全自动生长曲线分析仪在37°C下连续振荡培养21小时,每间隔一定时间自动测量OD600值,动态记录细菌生长曲线。通过比较野生型、cfa突变体及阳性对照dps突变体(已知对H₂O₂敏感)的生长延迟,评估CFA修饰对氧化应激抵抗的贡献。例如,突变体在H₂O₂存在下生长延迟延长至8.5小时,而野生型仅为5.5小时,表明CFA缺失加剧了氧化损伤。
实验结果
研究证实CFA缺失导致沙门氏菌对极端酸应激、氧化应激和质子动力势(PMF)破坏的敏感性显著增加。首先,cfa突变体在pH 3.0的极端酸条件下存活率大幅降低(野生型在3小时存活率显著高于突变体),表明CFA通过减少膜质子渗透性,帮助维持细胞内pH稳态。CFA修饰是沙门氏菌应对宿主内外环境压力的重要适应机制。CFA增强沙门氏菌在巨噬细胞内的生存能力,动物实验明确显示,cfa突变体在小鼠模型中毒力减弱。单独感染实验中,突变体感染的小鼠生存率高于野生型。阐明了CFA修饰的作用:CFA合酶(由cfa基因编码)在稳定期被诱导表达,通过将S-腺苷甲硫氨酸的亚甲基基团转移至膜不饱和脂肪酸,形成环丙烷环,从而增强膜脂的堆积密度和稳定性。这种修饰不仅降低质子泄漏、维持PMF,还提升膜对氧化剂的屏障功能。
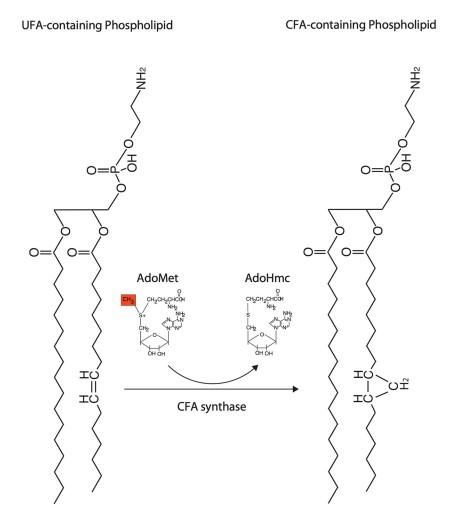
图1、环丙烷脂肪酸的生物合成,细胞膜中的磷脂作为反应底物。S-腺苷甲硫氨酸(AdoMet)中的亚甲基被转移至磷脂中不饱和脂肪酸的双键上,在烷基链上形成环丙烷环。催化该反应的细胞酶为依赖S-腺苷甲硫氨酸的甲基转移酶,即CFA合酶。
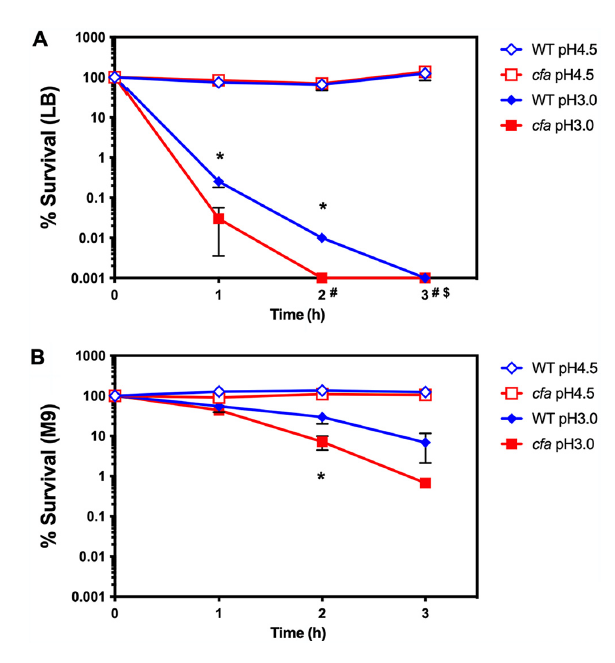
图2、环丙烷脂肪酸在极酸pH下的存活中必不可少。稳定期S.Typhimurium菌株在LB培养基(A)或M9培养基(含0.4%葡萄糖)(B)中培养,稀释1:1000接种至新鲜培养基,pH调至4.5(野生型,蓝色空菱形;cfa突变株,红色空方形)或pH 3.0(野生型,蓝色实菱形;cfa突变株,红色实方形)。在酸冲击处理后0、1、2和3小时测定存活CFU。$野生型和#cfa突变株在这些时间点低于理论检测下限。使用Student’s t检验比较野生型与突变株,*表示P<0.05。
相关新闻推荐
1、恶臭假单胞菌NCIMB 9866中PCMH和PHBDD在2,4-二甲苯降解中的关键作用(一)
2、细菌如何通过"慢生长"与"快生长"策略适应环境变化——以大肠杆菌为例的机制解析