嗜果糖特性
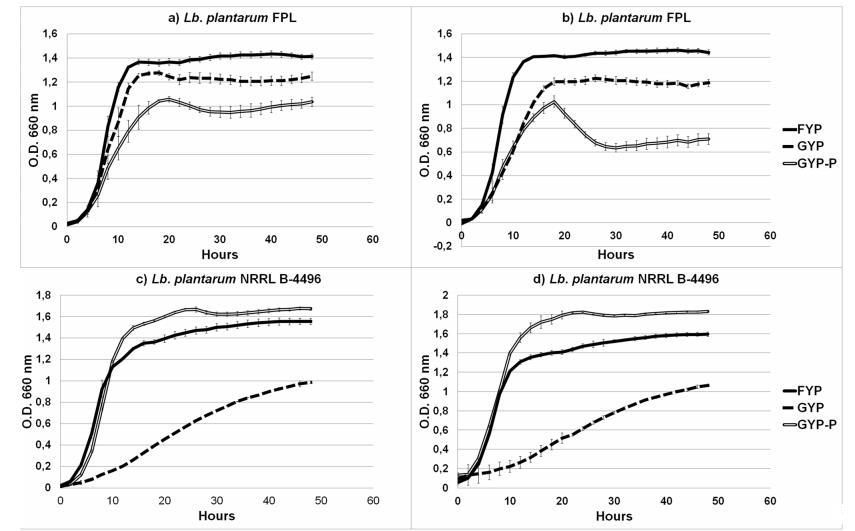
图3植物乳杆菌FPL在(a)有氧条件、(b)无氧条件及植物乳杆菌NRRL B-4496在(c)有氧条件、(d)无氧条件下不同碳源(FYP、GYP、GYP-P)中的生长曲线
所有测试的碳源都导致所用细菌的大量生长(即最终OD 660>0.9)。本研究中分离的菌株显示出非常相似的生长曲线;因此,图3仅显示了一个代表性菌株。生长参数显示在表3中。
表3 生长参数
| 菌株 | 培养基 | 生长条件 | 最大比生长速率 (小时⁻¹) | 延滞时间 (小时) | 倍增时间 (小时) | 最小 OD (中值滤波数据) | 最大 OD | 均方根误差 (RMSE) | R² |
|---|---|---|---|---|---|---|---|---|---|
| Lb. plantarum FPL | FYP (果糖) | 有氧 | 0.5822 | 0.0305 | 5.0423 | 0.0063 | 1.4630 | 0.0299 | 0.9991 |
| Lb. plantarum FPL | GYP (葡萄糖) | 有氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0125 | 0.9970 |
| Lb. plantarum FPL | GYP-P (葡萄糖+丙酮酸) | 有氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0235 | 0.9990 |
| Lb. plantarum FPL | FYP (果糖) | 厌氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0181 | 0.99 |
| Lb. plantarum NRRL B-4496 | FYP (果糖) | 有氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0075 | 0.9836 |
| Lb. plantarum NRRL B-4496 | FYP (果糖) | 厌氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0184 | 0.0975 |
| Lb. plantarum FPL | GYP (葡萄糖) | 厌氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0122 | 0.0185 |
| Lb. plantarum FPL | GYP-P (葡萄糖+丙酮酸) | 厌氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0531 | 0.0975 |
| Lb. plantarum NRRL B-4496 | GYP (葡萄糖) | 有氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0228 | 0.0995 |
| Lb. plantarum NRRL B-4496 | GYP-P (葡萄糖+丙酮酸) | 有氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0127 | 0.0986 |
| Lb. plantarum NRRL B-4496 | GYP (葡萄糖) | 厌氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0177 | 1.1987 |
| Lb. plantarum NRRL B-4496 | GYP-P (葡萄糖+丙酮酸) | 厌氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.1327 | 0.0063 |
| Lb. plantarum FPL | MRS (高糖) | 有氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0160 | 0.0937 |
| Lb. plantarum FPL | MRS (高糖) | 厌氧 | 0.0305 | 0.0302 | 8 | 0.0160 | 1.0637 | 0.0220 | 0.0220 |
菌株之间的另一个显著差异是在补充丙酮酸的培养基上的生长。丙酮酸以及氧气、柠檬酸盐和果糖可以被LAB用作外部电子受体,因此Endo等人在他们的实验中用它来展示嗜果糖乳酸菌的特征特性。我们的观察表明,丙酮酸也刺激植物乳杆菌FPL在葡萄糖上的生长,尤其是在有氧条件下。众所周知,植物乳杆菌在有氧条件下生长更好,而且,它能够利用氧气作为底物。此外,在氧气存在下,负责消耗糖的基因的表达增加。连续地,在有氧和厌氧条件下,参考菌株NRRL-4496在丙酮酸培养基上的生长都比我们的FPL菌株慢。这可以解释为NRRL-4496菌株不表现出嗜果糖特性。此外,最新的文章报道植物乳杆菌物种可以在各种代谢途径中利用丙酮酸。
尽管菌株NRRL B-4496在果糖上生长,但它不能在含有高浓度葡萄糖和果糖的培养基上生长。参考菌株没有发展出能够在富含糖的环境中生存的机制。已描述的FLAB的生长曲线与植物乳杆菌FPL相比更低。本文进行的实验表明,MALDI-TOF和Bioscreen C有助于快速筛选嗜果糖乳酸菌。
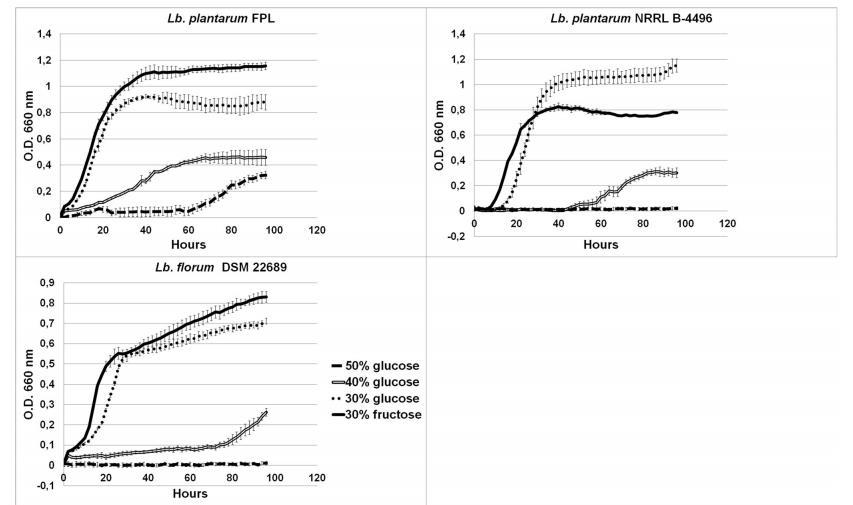
图4植物乳杆菌FPL、植物乳杆菌NRRL-4496、花乳杆菌DSM在不同高糖浓度培养基(葡萄糖或果糖含量为30%、40%和50%)需氧条件下的生长曲线
此外,新描述的菌株可以在含有50%(w/v)果糖的培养基上生长;除早金莲果杆菌外,其他FLAB耐受40%(w/v)的果糖含量。图4中的生长曲线显示了对高糖浓度生长的适应。在植物乳杆菌FPL菌株的情况下,嗜果糖性再次可见。该菌株在含有30%果糖的培养基上生长最好,其次是含有30%葡萄糖的培养基。在含有40%葡萄糖的培养基上可以看到缓慢的生长速率,甚至在含有50%葡萄糖的培养基上生长延迟。这种渗透压耐受性很高,因为通常细菌和酵母耐受高达50%(w/v)的糖。
在含有20、30、40、45和50%(w/v)葡萄糖或果糖的MRS和FYP培养基中,通过管底存在大量生物质证明了高糖浓度的耐受性。在含有20%和30%葡萄糖或果糖浓度的肉汤中孵育24小时后,细菌生长可见为生物质。48小时后,在含有40%、45%和50%糖浓度的培养基中可见生长;除了管底的生物质外,还有明显的浑浊。在果糖和葡萄糖上的生长差异不显著。在含有CaCO3的MRS和FYP琼脂上,菌落周围出现透明圈,表明产生酸。通过HPLC检测到上清液中有3.4%的乳酸。测试的细菌从葡萄糖产生气体,并且过氧化氢酶阴性。HPLC检测到培养的植物乳杆菌FPL菌株从果糖产生甘油;未检测到多元醇。此外,HPLC证实该菌株利用了两种碳源,但首先利用果糖。
许多FLAB以及植物乳杆菌FPL都是使用含有30%果糖的FYP分离的。还注意到在FYP上培养的分离株比在含有果糖的MRS上多。这证实了FYP是一种专门的培养基,其中高浓度的果糖选择FLAB,同时抑制其他细菌的生长。商业培养基中缺乏果糖解释了为什么嗜果糖细菌没有被更早地鉴定出来。植物乳杆菌菌株在各种环境中广泛分布,这可能是由于乳酸菌中最大的基因组之一。通常,乳杆菌具有相对大量的转运和调控基因以及糖转运和利用基因。
在植物乳杆菌WCFS1的基因组中,发现了30个糖转运系统,这解释了为什么该物种可以在多种碳源上生长。在植物乳杆菌中表达最多的基因中,除了管家基因外,还有糖酵解途径的基因和许多编码参与戊糖和己糖利用的酶的基因。全基因组测序表明,潜在高表达基因包括大量的磷酸转移酶系统,尤其是果糖和甘露糖PTS系统。这种灵活性可以解释植物乳杆菌FPL菌株嗜果糖特性的出现。此外,在本文中,我们描述了偏好果糖作为生长来源、具有高糖浓度抗性的菌株。如果扩展的植物乳杆菌基因组允许适应富含果糖的环境,是新FPL菌株嗜果糖特性的原因,那么这与FLAB组嗜果糖特征的起源是对立的。FLAB组对果糖的适应是由于缺乏adhE基因;在植物乳杆菌FPL的情况下,嗜果糖行为的机制有不同的基础(图5)。
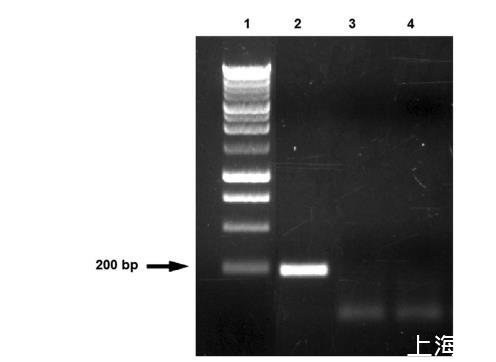
图5采用adhE引物获得的PCR扩增产物。第1泳道为1 kb Ladder Perfect Plus分子量标准(Eurx公司,格但斯克,波兰);第2泳道为Lb.florum DSM 22689的扩增产物;第3泳道显示Lb.plantarum FPL的扩增产物;第4泳道为Lb.plantarum NRRL B-4496
当然,这部分是由基因组中负责糖代谢和转运的大量基因解释的,但问题仍然是为什么与植物乳杆菌NRRL相比,该菌株更喜欢果糖而不是葡萄糖。植物乳杆菌在蜜露中的存在必须对蚜虫、蜜蜂或蚂蚁的生态系统产生影响。植物乳杆菌栖息在蚜虫和以蜜露为食的昆虫的消化道中也是非常可能的。生活在花蜜中的酵母增加了传粉媒介的访问次数,而花蜜中某些细菌(塔斯马尼亚欧文氏菌、库克乳杆菌、落新妇曲霉)的存在通过改变花蜜的化学成分来驱赶昆虫。应该研究更多来自波兰的蜜露样本,以确认植物乳杆菌对该栖息地的定殖及其对昆虫的影响。
结论
这项工作的主要目的是分离波兰蜜露中的嗜果糖乳酸菌,以了解来自温带气候地区蜜露中物种的变异性。我们的工作首次表明,来自欧洲温带气候的蜜露可能是一个有前景的新嗜果糖乳酸菌来源。据我们所知,选定的植物乳杆菌FPL菌株是第一个被描述为具有嗜果糖行为的植物乳杆菌菌株。植物乳杆菌在蜜露中的存在必须对蚜虫、蜜蜂或蚂蚁的生态系统产生影响。可以得出结论,植物乳杆菌FPL是一种共生细菌,栖息于褐软蚧的胃肠道中,并参与糖代谢。
相关新闻推荐
3、钙磷酸盐与PMMA骨水泥中纳米颗粒对抗耐药菌株的潜力与机制探讨(一)