ISP头部结构域参与与Qo位点各种占据物形成复合物。这些包括(i)启动分支反应的QH2和ISPox(2Fe-2S簇氧化)之间的酶-底物复合物(ES-复合物);(ii)ISPH(簇还原)和Q之间的酶-产物(EP-)复合物;以及(iii)ISPH和抑制剂之间的复合物,以UHDBT和stigmatellin为例。(iv)还解析了不同类型抑制剂的结构,以myxothiazol和MOA型抑制剂为例,它们结合在Qo位点更深处,不与ISP相互作用。涉及ISPH的复合物ii和iii可以通过它们的EPR光谱进行测定,因为还原的簇是顺磁性的,相互作用会在光谱中诱导出一条新线。在此基础上,可以确定参与这些复合物形成的物理化学性质,如SI附录中详述。特别是,因为复合物的形成将还原形式(ISPH)拉出氧化还原混合物,在每种情况下,氧化还原电位Em都会升高,其程度提供了结合强度DGapp的度量。底物结合形成ES-复合物也可以被测定,但由于该状态是亚稳态的,只能通过动力学研究。在每种复合物中,结合能的主要贡献来自ISP和位点占据物之间形成氢键。通过Nδ配位簇的组氨酸,当涉及ISPox时可以作为氢受体(-N-HO-),或者当涉及ISPH时可以作为氢供体(-NεH-O=)。
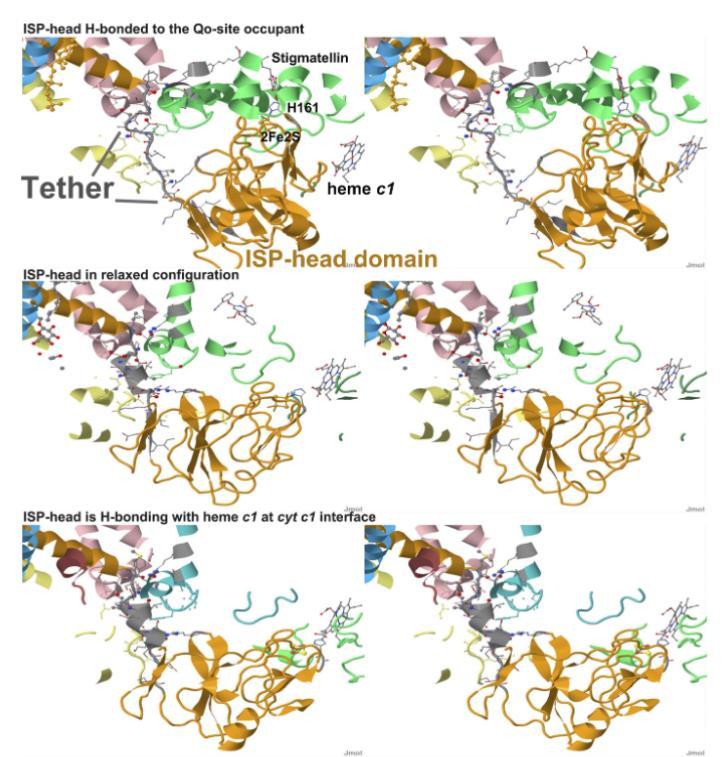
图6展示了ISP外域三种构象中弹簧加载机制的作用示意图。(上图)ISP与细胞色素b界面对接,通过氢键与Qo位点占据物[蛋白质数据库(PDB)编号3H1K,禽类复合物,结合了耻垢素]结合。主要特征已标注。(中图)ISP处于松弛构象[PDB编号3L71,禽类复合物,结合了唑氧菌素(作用机制抑制剂)]。(下图)ISP与细胞色素c1界面的血红素c1形成氢键(PDB编号1BE3,牛类复合物,未显示Qo位点占据物)。在松弛构象(中图)中,簇配体(牛类中的H161)未形成氢键接触,因此系链区域未受拉伸。
当H161与Qo位点占据物形成氢键时(上图),系链因外力作用(主要来自与占据物的氢键强度)而被拉伸,导致系链延长并旋转头部。螺旋区域被拉伸至伸展构象,从而导致多个跨跨间氢键断裂。当H161与血红素c1形成氢键时,位移幅度显著减小,系链区域仍保持与松弛态相似的氢键构型。SI附录图S6和表S6详细展示了Qo位点对接(耻垢素)与松弛构象(唑氧菌素)之间氢键变化的细节。链的颜色标注如下:链c(细胞色素b)为粉色;链d(细胞色素c1)为黄色;链e(ISP)为橙色;链j(7.2-kDa蛋白)为青色;链p(细胞色素b)为绿色(或在1BE3中为蓝绿色);链q(细胞色素c1)为橄榄绿(或在1BE3中为浅绿色)。灰色区域的卡通颜色由 CPK 的C原子或C-C键着色。邻近链7埃范围内的原子显示为半径0.2埃的填充球体,3.5埃范围内的原子显示为半径0.4埃的填充球体。选定原子按原子类型 CPK 着色。三个结构均以相同取向展示蛋白质支架,因此主要差异源于ISP锚定链和外在结构域的构象变化。交叉眼视立体图像
当形成复合物时(17,27-29,40),结构显示ISP头部与cyt b紧密堆积在界面上。否则,ISP头部旋转离开,组氨酸不形成氢键(图6)。这些不同的位置与连接外在结构域与其锚定螺旋的系链跨膜区的变化相关。当ISP与Qo位点占据物结合时,系链被拉成伸展构型,但当不结合时,系链塌陷成螺旋构型。从这些以及突变株中的类似研究(30,41,42)中,一个可能的机制变得显而易见(39)。在正常的复合物形成中,结合与一个约束力相平衡,主要来自螺旋中的氢键,在伸展过程中被破坏。当插入额外的残基以引入松弛,或者当通过将残基替换为螺旋破坏者(甘氨酸或脯氨酸)来阻止螺旋构型时,系链可以以较少的努力伸展,并且稳定复合物的氢键的表观强度DG增加。当系链被如此修饰以至于弹簧失去拉力时,ΔGappKd的值等于平衡力的损失。SI附录详细讨论了这一假设的物理化学基础。
类似的平衡力将在ES-复合物的形成中起作用,这些力与目前的讨论更相关,因为它启动了前向电子转移。头部结构域中与cyt b界面处的突变可能通过空间位阻抑制ES-复合物的形成,通过不利的VDW相互作用减少来自氢键的结合能,从而改变弹簧加载的平衡。通过限制ES-复合物的形成,此类突变,包括isp-1(qm150),将限制Qo位点的输入通量,从而限制SQo的占有率,而不影响其去除的反应。这种降低的占有率将降低ROS的产生,可能有助于长寿相关的效应。此外,减少的通量将传播到细胞的能量代谢,降低质子梯度,从而降低ATP合酶的电位,导致实质性的代谢和生理效应,这可以合理地解释许多多效性表型。然后,所有来自系链突变的抑制子功能都可以通过弹簧张力的缓解来轻松解释,使得ES-复合物更容易形成,输入通量得以恢复。来自早期突变研究的额外支持以及弹簧加载在控制和门控中的可能作用在SI附录中讨论。
在人类中,据报道,细胞色素b的特定突变与严重的心肌病和运动不耐受患者有关。将人类对应突变引入酵母 cyt b 会导致呼吸缺陷 (45)。在 Rip1 的系链区域内或 cyt b 中靠近 Rip1 系链的区域,鉴定出许多来自 cyt b S152P 和 G291D 突变(两者都位于 ISP 的对接界面)的呼吸缺陷表型的抑制子 (45)。与我们的发现类似,这些抑制子中有许多位于系链的丙氨酸残基中,这可能对系链区域的灵活性很重要。有趣的是,A90T 突变是 cyt b G291D 呼吸缺陷的较强抑制子之一,我们的数据表明,该突变 (sea4, A149T) 是我们在线虫筛选中鉴定的最有效的抑制子。总的来说,这些发现强调了系链区域在调节复合物 III 内 ISP 与 cyt b 相互作用中的重要性。
在这项研究中,我们已经证明,秀丽隐杆线虫线粒体复合物 III 的 Rieske 铁硫蛋白中突变的功能后果和相互作用,在高度进化分化的单细胞模式生物酿酒酵母中得到了复制。我们的数据提供了更多证据,表明系链区域内的基因内突变可以挽救不同生物体的呼吸缺陷。据我们所知,我们首次提供了一个讨论这些效应背后机制特征的框架,这带来了对复合物 III 在介导代谢速率、胚胎和胚胎后致死率、发育迟缓、排便周期、繁殖力、应激抵抗和寿命方面作用的分子理解。
意义
线粒体功能对健康和长寿至关重要。呼吸链中高度保守的Rieske铁硫亚基(ISP-1)的突变导致秀丽隐杆线虫出现多效性表型,包括发育迟缓和寿命延长。我们在ISP-1的一个保守的6个氨基酸系链区域中发现了基因内突变。这些抑制子能够抑制与isp-1(qm150)突变相关的所有表型。我们进一步证明,这种突变/抑制子关系在酵母复合物III的Rieske铁硫蛋白(Rip1)中是保守的。这些发现提供了对该蛋白质结构和功能保守特征的见解,并使我们能够提出一种独特的“弹簧加载”机制来解释这些效应,并得到经验物理化学数据的支持。
相关新闻推荐
1、Meta分析方法用于牛结节性皮肤病免疫抗体监测——讨论、结论
2、基于微生物生长曲线分析仪评价枯草三十七肽联合万古霉素对VRE的协同抗菌效应
3、驴用微生态制剂干酪乳酸杆菌筛选、生理生化鉴定及生长条件(二)