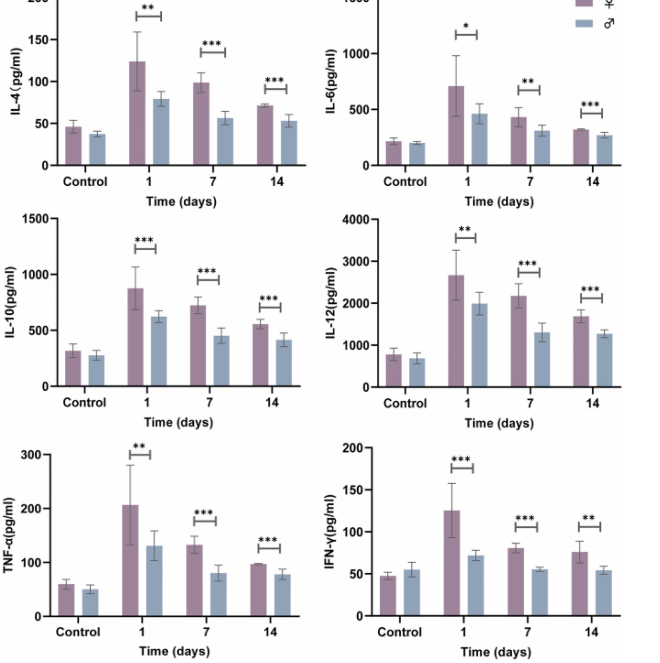
图2、雌性小鼠在法氏诺卡菌IFM 10152感染后表现出更高的细胞因子产生。雌性小鼠(n=32)和雄性小鼠(n=32)经鼻内感染1×10^7 CFU(50µl),分别在感染后1、7和14天,或在第1天感染50µl PBS,并通过ELISA测量肺上清液中的细胞因子水平(IL-4、IL-6、IL-10、IL-12、TNF-α和IFN-γ)。线图显示均值与标准误。数据来自2次独立实验。
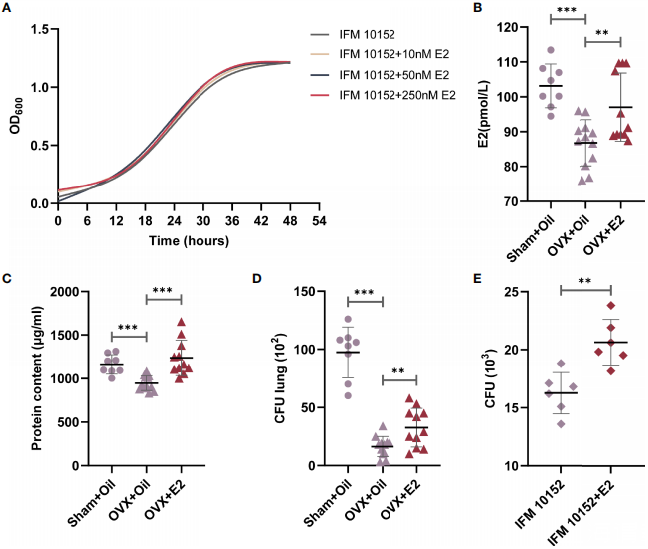
图3、E2补充增加去卵巢雌性小鼠对法氏诺卡菌IFM 10152肺部感染的易感性。(A)法氏诺卡菌IFM 10152在含有10 nM、50 nM和250 nM E2的BHI中培养48小时的生长曲线。(B–D)雌性小鼠在攻击法氏诺卡菌IFM 10152前接受假卵巢切除加芝麻油(n=8)、卵巢切除加芝麻油(n=12)或卵巢切除加E2(n=12)处理。(B)血清中E2水平。(C)BALF中蛋白含量。(D)肺组织匀浆中的细菌载量。(E)用或不用E2处理的肺泡巨噬细胞中的细菌存活情况。线显示均值与SEM。数据来自2次独立实验。
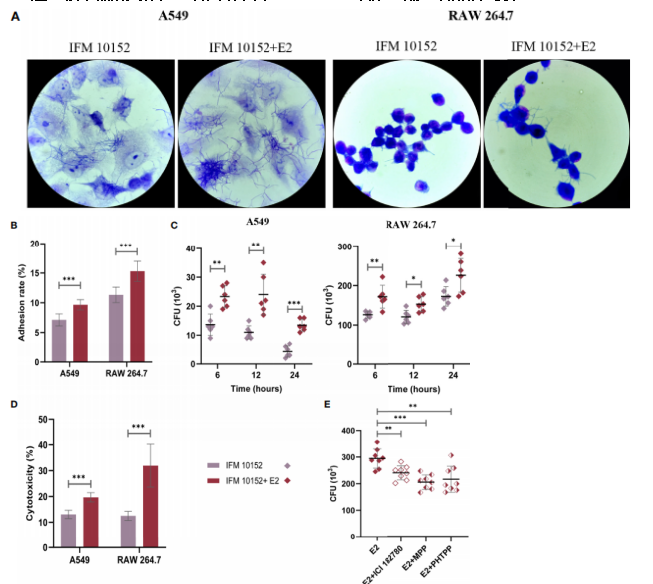
图4、E2通过ERα和ERβ信号促进法氏诺卡菌IFM 10152对宿主细胞的粘附和侵袭。(A–E)A549和RAW 264.7细胞在诺卡菌感染前用或不用50 nM E2处理16–18小时。感染1小时后细菌菌株对A549(左)和RAW 264.7(右)细胞的电子显微镜观察(A)和粘附率(B)。(C)感染后6、12和24小时细菌菌株对A549(左)和RAW 264.7(右)细胞的侵袭。(D)感染8小时后法氏诺卡菌IFM 10152对A549和RAW 264.7细胞的细胞毒性。(E)用ICI 182780、MPP或PHTPP处理的RAW 264.7细胞中的细菌存活情况。线显示均值与SEM。数据来自3次独立实验。
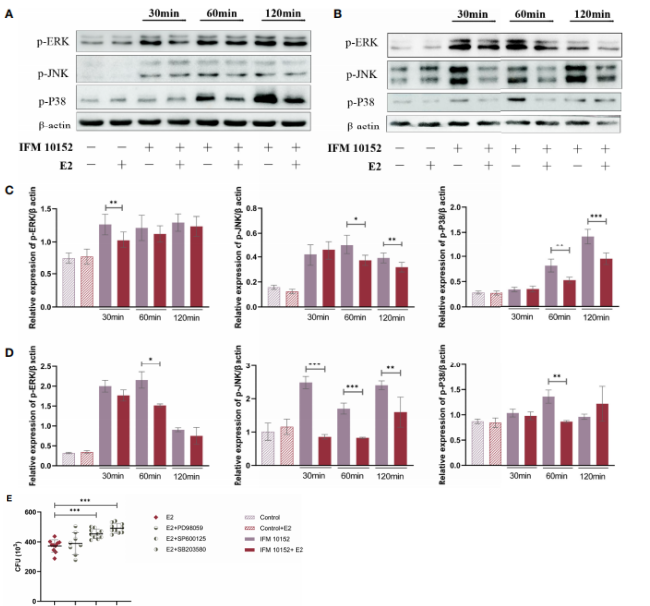
图5、E2通过下调MAPK信号通路促进细菌存活。A549(A)和RAW 264.7(B)细胞在诺卡菌感染前用或不用50 nM E2处理16–18小时。感染后30、60和120分钟ERK(p-ERK)、JNK(p-JNK)和p38(p-p38)磷酸化水平的Western Blot分析。通过Image J分析A549(C)和RAW 264.7(D)细胞中每种蛋白的相对表达量。(E)用20 mM PD 98059、20 mM SP 600125或20 mM SB 203580处理的RAW 264.7细胞中的细菌存活情况。线显示均值与SEM。数据来自3次独立实验。
总结
雄性通常比雌性更容易感染诺卡氏菌,雄性与雌性比例为2,临床疾病更高。17β-雌二醇在实验中通过抑制巴西猪笼草的生长来影响基于性别的间隙,但其潜在机制尚未完全阐明。然而在本研究中,我们报告了与雄性小鼠相比,感染法氏奈瑟菌IFM 10152的雌性小鼠的严重程度增加,死亡率增加、肺部细菌负荷升高和肺部炎症反应过度,这在补充E2的卵巢切除雌性小鼠中被模拟。
同样,在E2处理的宿主细胞中,细菌载量的压倒性增加也很明显,这与通过结合雌激素受体下调MAPK通路的磷酸化水平有关。我们得出的结论是,尽管男性诺卡氏菌感染的临床病例较多,但雌激素促进了细菌的存活,从而导致女性炎症加重。研究数据强调,在未来的诺卡氏菌研究中需要包括并单独分析两性,以了解免疫反应和疾病发病机制的性别差异。
Bioscreen设备在本研究中起到了关键的排除性验证工具作用,利用Bioscreen进行自动化、连续、定量的生长监测,避免了人工取样误差,提高了数据可靠性,有力支持了“E2的作用是宿主介导而非细菌自主”的核心论点。通过Bioscreen获得的高时间分辨率生长曲线,研究者明确发现——无论E2浓度如何变化,N.farcinica的生长动力学均未发生显著改变。这表明E2并非通过直接影响细菌增殖来加重感染。
相关新闻推荐
1、白星花金龟幼虫新鲜虫粪微生物菌群分离与鉴定——摘要、材料与方法
3、水稻黄单胞菌噬菌体最佳MOI (A)、一步生长曲线、耐受性测定及基因组分析(二)