乳酸乳球菌BirA与bioY基因的结合
相对于近亲猪链球菌的情况,由于存在birA和bioY的重复基因,bio基因座的基因组背景复杂(图8A)。在bioY1启动子区域前检测到推定的BirA1 DNA结合位点(ACA GTT AAC CTA AAT TTG ATT TTA GGG TTA CTG T)(图8B)。根据使用启动子预测中性网络程序预测的转录起始位点“T”的位置,BirA1结合位点似乎与“-10”到“-35”启动子区域重叠(图8B),表明bioY1的转录可能受乳酸乳球菌中BirA1负调控。
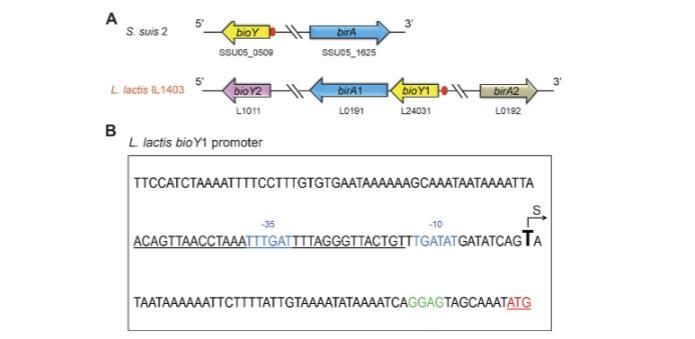
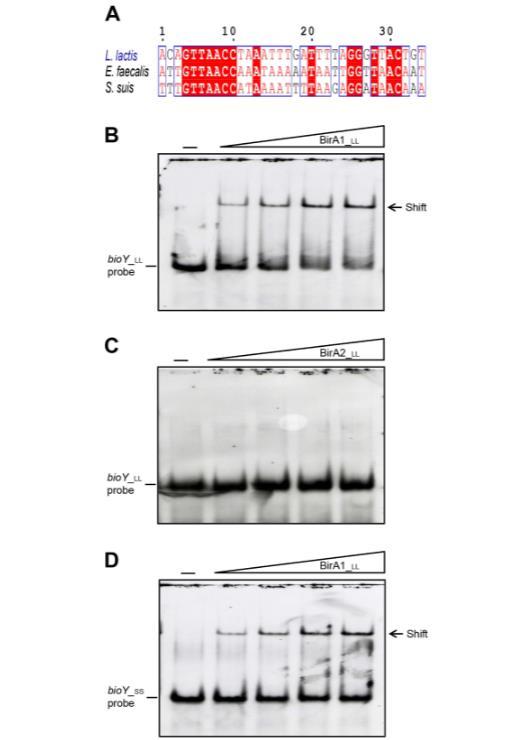
为了测试预测的BirA位点功能(图8A和9A),进行了电泳迁移率变动分析(EMSA),使用纯化的BirA1_LL和BirA2_LL(图9B-D)。EMSA证实BirA1_LL以剂量依赖性方式有效结合bioY1探针(图9B),而缺乏推定的N端DNA结合基序的BirA2_LL不结合探针(图2和9C)。有趣的是,猪链球菌bioY(bioY_ss)启动子能够与乳酸乳球菌BirA1_LL相互作用(图9D)。在猪链球菌BirA的情况下也发现了类似情况,它也能结合乳酸乳球菌bioY启动子。因此,我们预期猪链球菌和乳酸乳球菌的bioY和BirA之间存在串扰。鉴于BirA1和bioY1启动子之间发生相互作用,而bioY2缺乏预测的BirA1结合位点,探究这种调控机制与生物素同化的生理相关性可能很有意义。
乳酸乳球菌转运H-生物素的能力
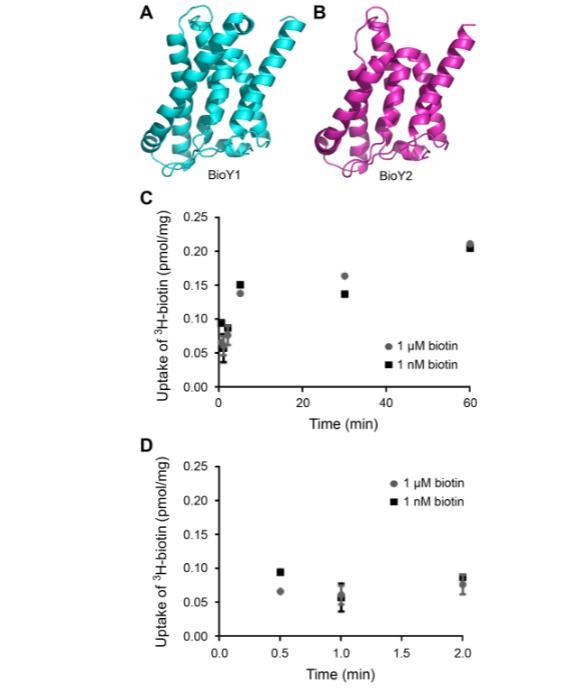
针对已解析的BioY结构的结构建模表明,BioY1_LL(图10A)和BioY2_LL(图10B)均由七个α-螺旋组成,并在整体构型上表现出明显相似性。鉴于乳酸乳球菌编码两个BioY转运蛋白,我们假设它能够进行强大的生物素转运。乳酸乳球菌培养物在限制生物素或过量生物素(1 nM或1μM)存在下生长4.5小时,离心沉淀并用PBS洗涤以去除游离生物素,用于3H-生物素转运测定(图10C,D)。在测量时间间隔内,生物素耗竭培养物(预测产生两种BioY转运蛋白)和生物素充足培养物(预测仅需单一转运蛋白)的3H-生物素相对转运相似。样品之间的微小差异仅在初始转运时间点观察到(图10D)。相反,在一分钟及以后,生物素的摄取/积累大致相等(图10C)。似乎测试的外源生物素增量并未显著增强生物素摄取活性。
结论
本文报告的数据表明益生菌细菌乳酸乳球菌中存在特定的生物素利用途径(图1),与其近亲动物病原体猪链球菌在birA和bioY基因座的冗余(和/或重复事件)上不同(图8)。乳酸乳球菌预计是生物素营养缺陷型,因为它缺乏完整的生物素生物合成途径。与猪链球菌中BirA调控bioY基因转录的生物素摄取调控机制不同,乳酸乳球菌有两个负责生物素摄取的BioY转运蛋白,和两个催化蛋白质生物素化的BPL酶(BirA1和BirA2_LL)。在两个bioY基因(bioY1_LL和bioY2_LL)中,bioY1_LL的转录似乎受BirA1_LL调控,而bioY2_LL的表达是组成型的(图1)。这种独特机制可能确保乳酸乳球菌具有响应环境和/或胃肠道中波动生物素水平的能力。
据我们所知,细菌生物素代谢的调控至少通过三种不同机制发生,分别以大肠杆菌BirA、农杆菌BioR和耻垢分枝杆菌BioQ为代表。调控的生物素转运以布鲁氏菌BioR22加上副球菌属12和猪链球菌通过BirA为例。乳酸乳球菌中两种不同功能性生物素蛋白连接酶的非典型出现增加了细菌生物素利用独特实例的广度(图1)。在弗朗西斯菌属中,我们也注意到两个不同BirA直系同源物的非典型存在。然而,我们未能检测到BioY转运蛋白的直系同源物,排除了调控生物素转运的可能性。与弗朗西斯菌属中双功能BirA蛋白在体外和体内与大肠杆菌bio操纵子发生串扰的情况不同,乳酸乳球菌BirA1不能调控大肠杆菌bio操纵子的表达(注意:蓝色是由于bio-lacZ融合的明显表达,图5)。值得注意的是,乳酸乳球菌BirA可以与猪链球菌bioY基因发生串扰(图9D),反之亦然。与大肠杆菌情况一致,在乳酸乳球菌基因组中检测到编码生物素受体蛋白的accB基因(L0187)(表S1)。乳酸乳球菌两个单独BirA同源物相对于单个大肠杆菌BirA的BPL活性降低(图6)可能表明BirA_LL的冗余是满足乳酸乳球菌蛋白质生物素化需求所必需的。乳酸乳球菌无法合成生物素以及胃肠道独特生态系统(充满各种营养元素和竞争细菌)可能选择了生物素清除途径的冗余,或者相反,基因组织可能只是细菌进化的遗迹。未来工作可以探索不同生物素条件下转运蛋白产生的调控以及转运蛋白效率的差异。