aw、pH值和抑菌剂等因素同样对微生物的生长产生极大影响。干燥条件是控制奶粉生产过程中阪崎克罗诺杆菌的重要参数。Lang等通过比较不同干燥处理后阪崎克罗诺杆菌的生长参数,证明底物的干燥速率和干燥后的最终aw是造成菌株不可逆损伤的重要条件,该研究为干燥后病原微生物的监控提供参考。类似地,Zhu等发现阪崎克罗诺杆菌不可在用乳酸、乙酸等有机酸酸化至pH 5的培养基中生长,此研究结果可为酸化奶粉研制提供理论支持。
另外,抑菌剂的存在也可对菌株的生长产生影响,很多已有研究都可以证明这一点。例如Shi Chao等采用Gompertz模型来定量描述柠檬醛或者百里醌对胰蛋白胨大豆肉汤培养基(tryptone soya broth,TSB)中阪崎克罗诺杆菌生长的影响,发现柠檬醛和百里醌均可以延长迟滞期,降低比生长速率。为加快抑菌物质在食品中的应用,确保环境因素成为食品安全的辅助工具,模型验证工作刻不容缓。模型验证的主要问题不在于模型对数据的拟合程度,而在于如何准确地模拟微生物对给定环境的响应。综上,利用环境因素降低奶粉贮藏、食用过程中阪崎克罗诺杆菌风险的研究尚处于初级阶段,建议继续加强阪崎克罗诺杆菌的风险建模研究,完善模型验证工作。
另外,菌株在食品工业环境或者食品加工过程中可能会受到一定的应激作用。对于受热应激(60.0℃、2 min)的阪崎克罗诺杆菌,Fang Ting等使用Huang模型拟合其在不同温度下的生长曲线。对温度与最大比生长速率的关系选择修正的Huang模型进行描述,数据显示热应激并没有引起最大比生长速率的显著变化。类似的研究也见于Yang等对受热应激、酸应激和干燥应激的阪崎克罗诺杆菌在室温下生长模型的构建,显著性分析表明迟滞期同样没有因为应激作用而发生显著变化。除了迟滞期和最大比生长速率,生理状态也可描述历史应激对微生物的生长作用。生理状态概念由Baranyi等提出,用来描述生长历史对迟滞期的影响。综上所述,虽然目前已经建立了历史应激对群体迟滞期影响的模型,但是关于历史应激与生理状态的关系尚待完善。
此外,虽然群体迟滞期的定义已经在微生物学中广泛使用,但应激条件下其与单细胞迟滞期的关系在阪崎克罗诺杆菌研究中鲜有阐述。单细胞生长模型的建立多采用比浊法,利用Bioscreen软件测量的浊度曲线估计微生物的生长参数是一种相对简单的方法,建议将来可结合此方法对低菌量阪崎克罗诺杆菌的生长进行建模。
2.1.2随机性模型
除了环境因素,微生物的初始污染水平及菌株变异性对其生长也会产生极大影响。据调查,实际流通食品中致病菌的含量通常较低,多数奶粉中阪崎克罗诺杆菌的污染水平小于1 CFU/100 g。此时可以用随机性模型描述较小微生物种群或单个细胞的生长行为。Foods等获得形状参数为5的极值II型(EVIIb)分布是描述阪崎克罗诺杆菌单细胞迟滞期分布的最佳模型,可为国际标准化组织对阪崎克罗诺杆菌标准检测方法的修订提供借鉴。此外,为探究历史应激对单细胞变异性的影响,Xu Yao等将单细胞的迟滞期假设为带有固定形状参数的移位Gamma分布,研究发现热应激使得阪崎克罗诺杆菌单细胞迟滞期的分布向右移动,单细胞迟滞期的变异性随恢复温度的降低而增加。在今后的研究中,考虑微生物的变异性是至关重要的,因为微生物风险评估的可信度是基于其估计最终风险所涉及参数的变异性和不确定性的能力。若奶粉中存在少量迟滞期短的非典型菌株,会缩短食品的保质期、增加消费者的健康风险。另外,为更加精准地估计风险,变异性应纳入剂量-效应模型。
综上所述,确定性模型的结果是由初始条件唯一确定的精确数值,通常用来表示群体微生物的生长。虽然传统模型很容易理解,但是它们没有考虑到菌株之间以及单个细胞之间的差异性,因此确定性模型不适用于描述低菌量或者单细胞水平下微生物的生长规律。随机性建模很好地弥补了传统预测微生物模型进行点估计的不足,该方法的输出结果很好地提供了关于微生物污染分布和超过特定临界水平的概率等信息。在模型的应用上,一级模型中,Gompertz模型因为简便易懂而得到广泛应用,Baranyi模型因为在生长研究中具有较好的拟合效果而被应用于估计最大比生长速率等参数。由于温度在食品生产消费链中波动最大,其成为阪崎克罗诺杆菌二级模型研究最多的因素。利用预测微生物学模型研究不同条件下阪崎克罗诺杆菌的生长可为实际食品病原微生物的监测和控制提供有效信息。不同环境条件影响阪崎克罗诺杆菌的生长模型及参数如表1所示。

表1部分不同条件下阪崎克罗诺杆菌的生长模型及参数
2.2失活模型
预测微生物学试图提供在各种环境条件下的微生物失活模型,例如温度、pH值、aw、抑菌剂和其他新型抑菌技术的作用等。因此,预测模型可以看作是栅栏技术的量化,失活模型及其参数可以反映不同控制方法对微生物的作用强度和效果,也可以反映微生物经历的不同应激条件对其失活的影响。
2.2.1环境因素对失活的作用
热处理是食品工业和家庭环境灭活微生物并确保良好产品稳定性的优选技术,因此关于阪崎克罗诺杆菌的热失活研究相对较多。温度对微生物失活的影响通常使用一级线性动力学方程获得的D值(在一定的处理环境中或一定的热力致死温度条件下某细菌中每杀死90%原有残存活菌数时所需要的时间)和Z值(灭菌时间减少到原来的1/10或在相同灭菌时间内杀灭99%的微生物所需升高的温度)表示。早在1996年,Nazarowec-White等指出阪崎克罗诺杆菌是肠杆菌科最耐热的成员之一,58℃时复水奶粉中阪崎克罗诺杆菌D值为5.45 min;而Breeuwer等发现有些阪崎克罗诺杆菌在58℃时D值范围为0.27~0.90 min。同一温度下不同研究中D值差异较大,表明菌株之间耐热性存在差异,但这并不妨碍温度对阪崎克罗诺杆菌的作用效果。从表2中由失活模型获得的参数可见,温度越高D值越小,奶粉在复水过程中适当提高环境温度可以有效提高杀菌效果。温度对阪崎克罗诺杆菌的失活研究可为奶粉制造商和婴幼儿护理人员提供奶粉复水温度的理论依据和数据支持。
除了热处理,近年来控制阪崎克罗诺杆菌的新技术也不断发展,利用非热技术控制病原微生物的繁殖逐步成为提高产品质量和保障食品安全的重要策略,而失活模型及其参数成为阐述新技术对微生物作用效果的重要依据。Lee和Osaili等的研究表明,低剂量的γ射线便可有效灭活奶粉中的阪崎克罗诺杆菌,因此可以进一步研究γ射线应用于企业生产的可行性。除了采用传统的线性失活模型表达微生物的失活状况外,非线性模型可以更加精确地阐述新技术对微生物的控制效果,例如Arroyo等采用Weibull模型、Adekunte等采用带有“肩带”的对数线性模型和Bigelow模型将阪崎克罗诺杆菌的失活数据拟合到对应模型以获得失活参数,从而证明了超声处理、高强度脉冲电场技术对菌株的灭活效果。除了动力学模型,随机性模型同样可以很好地对阪崎克罗诺杆菌的失活数据进行拟合。Koseki等建立了一个生存/死亡界面模型,该模型用于确定经高静水压处理后阪崎克罗诺杆菌的生长、失活行为。由于该模型是基于概率的方法所建立,其可以估计微生物达到预期失活效果时的失活条件,同时估计该条件下达预期效果的成功概率。上述模型的研究结果将有助于企业或者管理者同时考虑多个环境因素设定婴儿配方奶粉生产、加工标准。
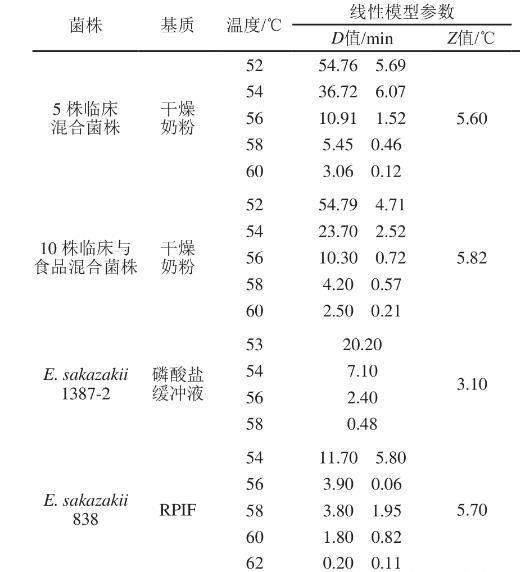
表2部分阪崎克罗诺杆菌热失活的线性模型及参数
相关新闻推荐
1、AgBr纳米颗粒抑制变形链球菌生长与防止生物膜形成,适用于义齿清洁行业