2结果
2.1重组质粒pUC19-△BspE的构建及鉴定
以牛种布鲁氏菌A19菌液作为模板,利用PCR反应对BspE基因上、下游同源臂进行扩增,结果显示,获得大小均为601bp的特异性条带(图2A),与预期相符;将BspE基因上、下游同源臂按1:1混合后,使用融合PCR方法进行扩增,结果显示,融合片段大小为1202bp(图2B),与预期相符;将融合片段与pUC19-SacB载体连接,得到重组质粒pUC19-△BspE,经PCR检测获得大小为1202 bp的特异性条带(图2C),测序鉴定结果与预期相符,表明重组质粒构建正确,可用于后续筛选BspE基因缺失株。
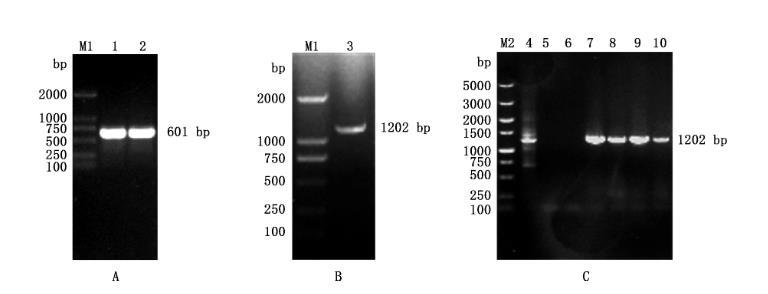
图2重组质粒pUC19-ABspE的构建及鉴定
2.2重组质粒pBB-BspE的构建及鉴定
以牛种布鲁氏菌A19菌液、pET-28a(+)和pBBR1MCS作为模板,分别对BspE、kana基因及pBB复制子基因进行PCR扩增,结果显示,分别获得大小为675、1229和1741bp的特异性条带(图3A~3C),与预期相符;将BspE基因片段与kana基因片段按1:1比例混合后,使用融合PCR方法进行扩增,结果显示,融合片段大小为1904 bp(图3D),与预期相符;将融合片段与pBB复制子片段连接,得到重组质粒pBB-BspE,经PCR检测获得大小为675bp的特异性条带(图3E),经测序鉴定其与预期相符,表明重组质粒构建正确,可用于后续筛选BspE基因回补株。
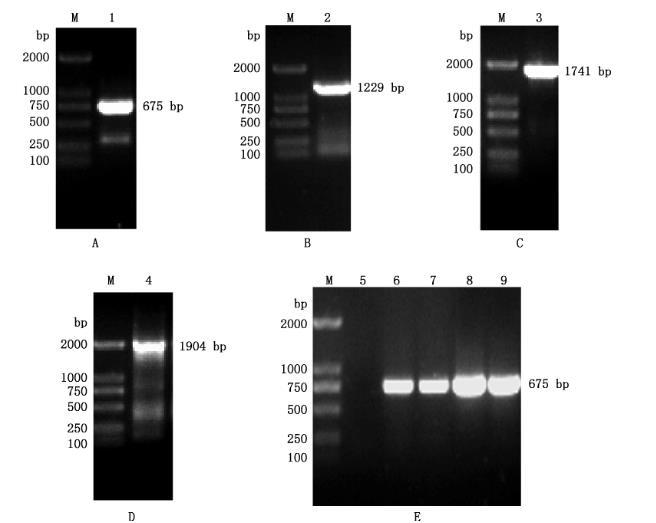
图3重组质粒pBB-BspE的构建及鉴定
2.3布鲁氏菌BspE基因缺失株筛选
选择阳性菌落进行PCR检测,结果显示,在样品5和7中扩增出大小为2060bp的特异性条带(图4),与预期相符,表明BspE基因缺失株A19ΔBspE构建成功。
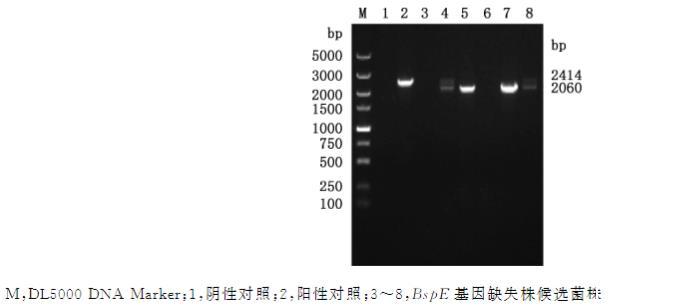
图4牛种布鲁氏菌BspE基因缺失株筛选
2.4布鲁氏菌BspE基因回补株筛选
选择阳性菌落进行Western blotting检测,结果显示,检测到大小约为14.59ku的BspE-flag条带(图5),表明BspE基因回补株A19C△BspE构建成功。
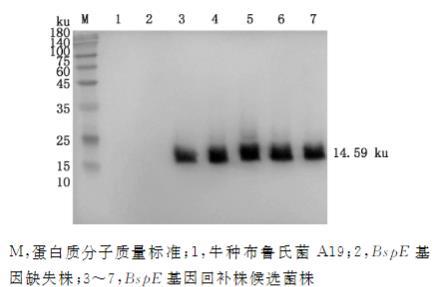
图5牛种布鲁氏菌BspE基因回补株筛选
2.5 BspE基因缺失对布鲁氏菌体外生长的影响
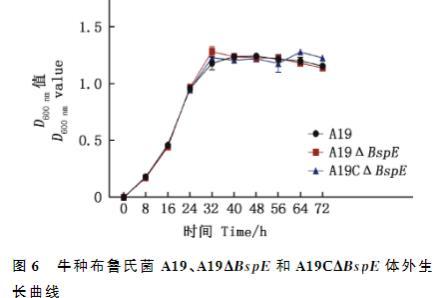
细菌生长曲线测定结果显示,与牛种布鲁氏菌A19和A19C△BspE相比,在24~40h测定时间内,A19△BspE呈上升趋势,56h后下降,但三者之间的生长变化并无显著差异(P>0.05);与牛种布鲁氏菌A19和A19△BspE相比,在40~56h测定时间内,A19C△BspE呈下降趋势,56h后上升,但三者之间的生长变化并没有显著差异(P>0.05);在所有测定的时间内,牛种布鲁氏菌A19、A19△BspE与A19C△BspE的生长趋势均没有显著差异(P>0.05,图6),说明BspE基因的敲除对布鲁氏菌的体外生长没有影响。
2.6 BspE基因缺失对布鲁氏菌胞内生存的影响
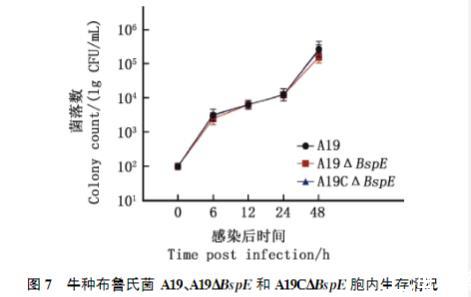
由图7可知,与牛种布鲁氏菌A19和A19C△BspE相比,在感染6和48h后,A19△BspE在RAW264.7细胞内生存能力呈下降趋势,但三者之间的胞内生存能力并无显著差异(P>0.05),均在8h达对数生长期,32h进入平台期;在感染0、6、12、24及48h后,A19△BspE与牛种布鲁氏菌A19、A19C△BspE的胞内生存能力均无显著差异(P>0.05),说明BspE基因的缺失对布鲁氏菌在胞内的存活情况没有影响。
2.7 BspE基因缺失对布鲁氏菌黏附能力的影响
细菌黏附试验结果显示,与牛种布鲁氏菌A19和A19C△BspE相比,A19△BspE对RAW264.7细胞的黏附能力没有显著差异(P>0.05,图8),说明BspE基因的敲除对布鲁氏菌在胞内的黏附能力没有影响。
图8牛种布鲁氏菌A19、A19ΔBspE和A19CΔBspE的黏附能力
2.8 BspE基因缺失对布鲁氏菌入侵RAW264.7细胞能力的影响
细菌入侵试验结果显示,与牛种布鲁氏菌A19和A19C△BspE相比,A19△BspE人侵RAW264.7细胞的能力没有显著差异(P>0.05,图9),说明BspE基因的敲除对布鲁氏菌入侵细胞的能力没有影响。
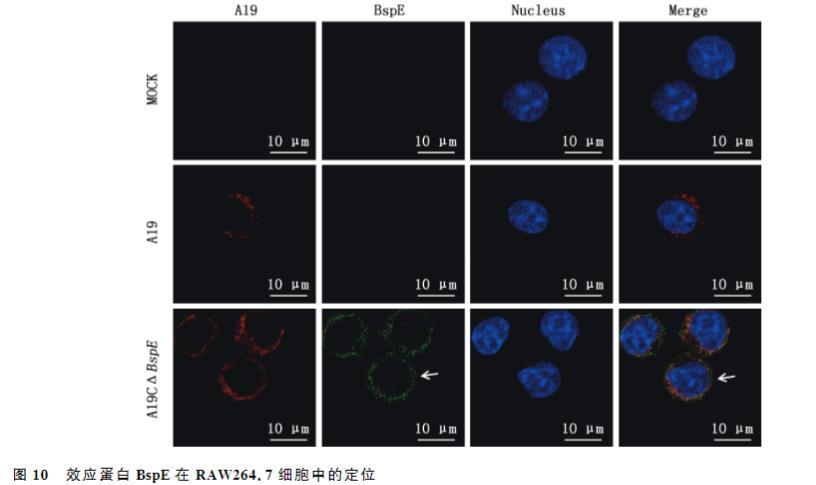
2.9效应蛋白BspE在小鼠RAW264.7细胞中的定位
由图10可知,在未感染的MOCK组中并没有出现红色荧光和绿色荧光;牛种布鲁氏菌A19感染组中只出现了红色荧光,没有出现绿色荧光;带flag标签的BspE蛋白的A19C△BspE感染组中出现了红色荧光和绿色荧光(箭头所示),且效应蛋白BspE大多分布于核周区域,表明效应蛋白BspE主要定位于感染细胞的核周区域。
3讨论
布鲁氏菌是一种胞内寄生的革兰阴性细菌,缺乏典型的外毒素、荚膜等毒力因子。布鲁氏菌毒力因子有助于细菌躲避机体免疫杀伤,从而侵入宿主细胞,完成胞内增殖并释放至胞外造成疾病。T4SS是布鲁氏菌重要的毒力因子,其可调节细菌效应蛋白,并转运至宿主细胞,有利于布鲁氏菌感染宿主细胞。
本研究利用同源重组结合SacB反向筛选技术成功获得了无痕缺失的牛种布鲁氏菌A19效应蛋白BspE基因缺失株A19△BspE。王玉飞等利用含蔗糖致死基因SacB的pEX18Gm质粒成功构建了缺失T4SS启动子区的羊种布鲁氏菌无痕敲除株。Tian等以SacB为负筛选标记,成功敲除了鸭疫里默氏杆菌CH-1的dps基因。
生长曲线结果显示,在所有测定的时间内,牛种布鲁氏菌A19、A19△BspE与A19C△BspE的生长趋势均没有显著差异,表明BspE基因的敲除不会影响布鲁氏菌的生长。这与以往研究者发现布鲁氏菌T4SS效应蛋白VceA、VceC、BPE043及BPE159等不影响布鲁氏菌的体外生长结果一致。
布鲁氏菌在宿主细胞内的增殖主要是在感染后24~48h。本研究比较了牛种布鲁氏菌A19、A19△BspE、A19C△BspE在小鼠RAW264.7细胞中的生存能力。结果显示,A19△BspE与牛种布鲁氏菌A19、A19C△BspE在RAW264.7巨噬细胞内的生存能力均没有显著差异,表明BspE基因的缺失不会影响布鲁氏菌在胞内的存活情况。这与徐朕宇等研究发现缺失牛种布鲁氏菌疫苗株A19效应子BtpA基因后不影响布鲁氏菌在小鼠RAW264.7细胞中的生存结论基本一致。
黏附对布鲁氏菌感染宿主细胞极为重要。本研究对牛种布鲁氏菌A19、A19△BspE、A19C△BspE黏附入侵RAW264.7细胞的能力进行了比较。结果显示,A19△BspE黏附和入侵RAW264.7细胞的能力与牛种布鲁氏菌A19、A19C△BspE均没有显著差异,表明BspE基因的缺失不影响布鲁氏菌黏附、入侵RAW264.7细胞的能力。这与邓肖玉等、陈蕾等发现牛种布鲁氏菌A19 virB启动子缺失和猪种布鲁氏菌S2入侵相关基因ialB缺失后不影响布鲁氏菌对RAW264.7细胞的黏附入侵能力的结论基本一致。上述结果表明布鲁氏菌的BspE可能在体外培养及体外感染细胞的过程中并不发挥对细菌黏附、入侵及胞内生存的调控作用,其重要的生物学功能需进一步探究。
IFA结果显示,在感染24h后,BspE主要分布于核周区域。Myeni等在HeLa细胞中转染HA-BspE表达质粒后,通过荧光共聚焦显微镜观察其主要在核周区域形成离散的囊泡,而本研究在构建的缺失菌株A19△BspE上回补BspE质粒,在菌体上进一步证实了BspE主要定位于感染细胞的核周区域。
综上所述,效应蛋白BspE的缺失不影响布鲁氏菌在RAW264.7细胞中的生存及黏附入侵细胞的能力,其在布鲁氏菌感染过程中的具体作用尚需进一步探究。由于BspE的缺失并不影响布鲁氏菌在体外及细胞感染过程中的生长,提示可针对BspE功能位点进行标记疫苗或鉴别诊断类产品的开发。本试验结果为进一步研究布鲁氏菌效应蛋白BspE的生物学功能及致病机制奠定基础。
4结论
本研究通过构建牛种布鲁氏菌效应蛋白BspE基因缺失株和回补株,发现效应蛋白BspE的缺失不影响布鲁氏菌在体外以及RAW264.7细胞中的生存、黏附、入侵能力,且效应蛋白BspE主要定位在感染细胞的核周区域。