2.6 YJ-22分离株免疫原性验证结果
利用YJ-22分离株制备灭活疫苗免疫兔子后,通过ELISA检测血清中特异的抗M.bovis IgG抗体。结果显示(表3):
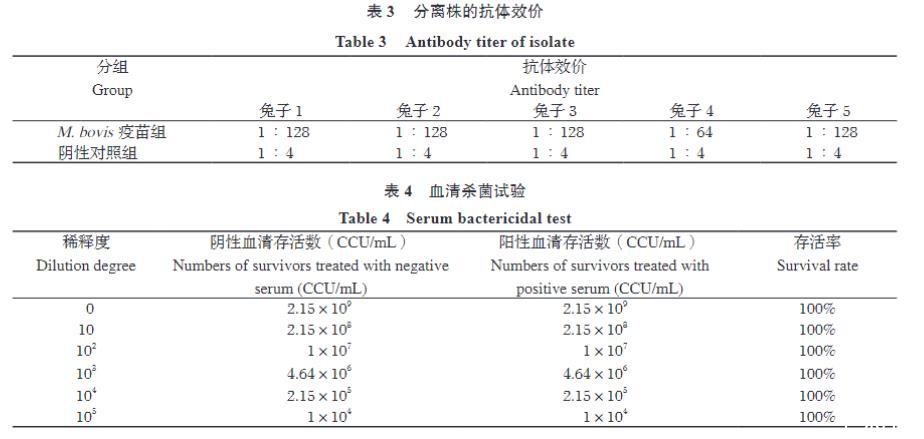
阴性对照组(注射生理盐水)的5只兔子,血清抗体效价均不高于1:4(均在1:4稀释度低于Cut-off值)。
M.bovis疫苗免疫组的5只兔子,二免后14天血清抗体效价均显著升高,其中4只达到≥1:128(终点效价为1:128),1只为1:64。所有免疫兔子的抗体效价均≥1:64。
这表明YJ-22灭活疫苗能有效刺激兔子产生高水平的特异性IgG抗体应答,证明该分离株具有强免疫原性。
2.7兔抗血清体外杀菌试验
为了初步评估高免兔血清是否具有杀菌活性,进行了体外杀菌试验。将不同稀释度的YJ-22菌液分别与经除菌灭活处理的阳性兔血清(来自效价≥1:64的免疫兔)或阴性兔血清混合孵育2小时后,测定存活的细菌数(CCU)。
结果(表4)显示,所有稀释度(10?~10??)的YJ-22菌液,经免疫兔阳性血清处理后的CCU与其相应的阴性血清处理后的CCU值相比,没有显著降低或升高。计算各稀释度的M.bovis存活率。结果显示,在测试的所有稀释度下,M.bovis存活率均接近或等于100%(表4)。这表明,在本试验条件下,虽然兔抗血清中含有高水平的特异性IgG抗体,但这些抗体在体外并无杀灭或抑制YJ-22生长的作用。
表中A、B代表各稀释度三次独立试验的平均CCU值(示例性,实际需填写或注明见图/下表)。本试验三次重复中,所有稀释度下,阳性血清处理组菌液在孵育2h后的CCU均与阴性血清处理组无显著差异,存活率计算结果均为100%。数据以平均值±标准差表示,无统计学差异(P>0.05)。_
*(注:这里展示的是结果表述方式,实际数据应以图或补充表格形式完整呈现三次试验结果及计算过程。表4核心信息为“所有稀释度下存活率=100%”)_
3讨论
牛支原体感染在养牛业中广泛存在,但由于其种类多,且牛源支原体如无乳支原体(M.agalactiae)与牛支原体在生化特性(如都不能利用精氨酸)及16S rRNA序列(>99%相似度)上高度相似,对其进行准确鉴定至关重要。本研究采用靶向M.bovis特异性表面脂蛋白P48基因的PCR方法进行鉴定,结合菌落形态特征(典型的“煎蛋样”外观)和生化鉴定结果(特别是利用乳糖和生化全套结果符合M.bovis特征),成功从患严重肺炎的犊牛肺组织中分离出一株致病性牛支原体YJ-22。测序证实其P48基因片段与国内外多个参考株(如MJ-1、DADC51、VK30)高度同源(>99.3%),进一步确认了菌株种属。
鉴于支原体基因组小(YJ-22同源株基因组约1 Mb)、代谢途径缺乏、营养需求苛刻,在体外实现高密度培养是其研究难点。我们研究了YJ-22在含血清PPLO培养基中的生长特性。其生长周期约为60小时,平台期出现在接种后18~45小时,此时菌体浓度最高可达10?CCU/mL。该平台期时间窗较长(约27小时),且菌体浓度较高,为YJ-22的规模化培养(如抗原制备、代谢组学分析等)提供了参考依据。选择合适的收获时间点(如平台期中期)可获取最大菌体量。值得注意的是,45小时后进入衰亡期,菌体浓度下降明显,提示收获时机的重要性。
为了评价YJ-22的致病性,我们采用连续3天气管注射攻毒的方式感染2~3月龄断奶犊牛。结果显示该攻毒模型成功复制了M.bovis肺炎的典型临床特征(发热、咳嗽、鼻腔分泌物)和病理损害(肺部大面积肉变、肝变、出血;组织学证实为伴有单核巨噬细胞浸润的支气管间质性肺炎),攻毒牛出现持续性排菌(鼻拭子阳性率达40-50%),肺部病变评分(9和11分)显著高于对照组(1和2分)。这证明YJ-22是一株具有较强致病力的牛支原体分离株。本研究采用连续攻毒策略,可能与单次大剂量攻毒相比,更接近自然感染过程(持续暴露),并有效诱导了肺部严重病变。研究结果为后续疫苗攻毒保护效力评价提供了一个有效的攻毒模型。
由于M.bovis对常规抗生素(特别是靶向细胞壁类)具有固有抗性,且易产生获得性耐药,疫苗防控是理想策略。然而,商品化疫苗的缺乏凸显了研发困境。利用YJ-22制备灭活疫苗免疫兔子,成功诱导了高水平的特异性IgG抗体应答(效价≥1:64),证实该菌株具有良好的免疫原性,其表达的抗原能被宿主免疫系统识别并产生强应答,这使其成为潜在的疫苗候选株(包括灭活疫苗组分或用于基因工程抗原筛选)。兔子体型较小、易于操作,是验证M.bovis分离株/抗原免疫原性的常用模型。
然而,免疫原性强并不意味着保护性免疫。体外杀菌试验揭示了重要发现:尽管高效价抗血清中含有大量特异IgG,但这些抗体在体外并不能杀灭或抑制YJ-22的生长(存活率始终为100%)。这提示:
1.抗体类型或靶点:这些抗体可能并非中和抗体。M.bovis存活的关键可能依赖于某些难以被抗体阻断的机制,如快速的抗原变异(逃避)、侵入宿主细胞、黏附后的定植防护等。产生的IgG可能主要识别非保护性的表面蛋白。
2.免疫保护机制:M.bovis感染的清除机制可能更依赖于细胞免疫(如巨噬细胞活化、T细胞介导的免疫)。体液抗体可能仅起辅助作用,如调理吞噬、阻断黏附(需进一步验证抗体能否抑制黏附)或通过Fc效应激活补体等(本试验灭活了补体)。需要设计黏附抑制试验、调理吞噬试验或添加外源补体的试验来深入评估抗体功能。
3.体外模型局限性:CCU法主要检测代谢活性,在无抗体介导效应分子(如补体、吞噬细胞)的单纯培养体系中,抗体直接作用于支原体本身的效果可能确实有限。
4.体内保护效力是关键:体外缺乏杀菌活性不等于体内缺乏保护作用。疫苗的保护效果最终需要在易感宿主(牛)中通过攻毒保护试验进行严格评价。
4结论
本研究成功分离并鉴定出一株具有强致病力的牛支原体YJ-22。该菌株能良好生长,在标准培养基中可获得较高浓度的菌液。利用该分离株建立的犊牛气管注射连续攻毒模型,能稳定复制出M.bovis肺炎的典型临床和病理特征。YJ-22对兔子具有强免疫原性,可诱导高水平的特异性IgG抗体应答,证明其含有有效的免疫原组分。然而,体外试验表明这些抗血清不具有直接杀菌作用,提示针对M.bovis的保护性免疫机制可能更为复杂。综上,YJ-22分离株作为一株具有明确致病性和免疫原性的地方流行株,为深入研究M.bovis的生物学特性、致病机制以及未来疫苗(尤其是以兔子模型筛选保护性抗原或验证免疫策略)的研发提供了重要的候选菌株和实验基础。后续研究应重点在牛模型中评估基于YJ-22的疫苗的保护效力,并深入探究其免疫保护机制(侧重细胞免疫和功能性抗体评价)。
相关新闻推荐
2、新发现BfmAK信号系统控制嗜麦芽寡养单胞菌生物膜形成(四)
3、自然发酵鲊肉粉中筛选乳酸菌和葡萄球菌生长曲线及生物学特性(三)