2.2鼠李糖乳杆菌Probio-M9连续传代过程中比较基因组学分析
2.2.1基因组信息本研究中不同代时菌株组装的基因组信息如表3所示,鼠李糖乳杆菌Probio-M9原始菌株基因组大小3.00 Mb,GC含量46.76%。传代过程中菌株的平均基因组大小为(3.00±0.002)Mb,GC含量(46.76±0.007)%。
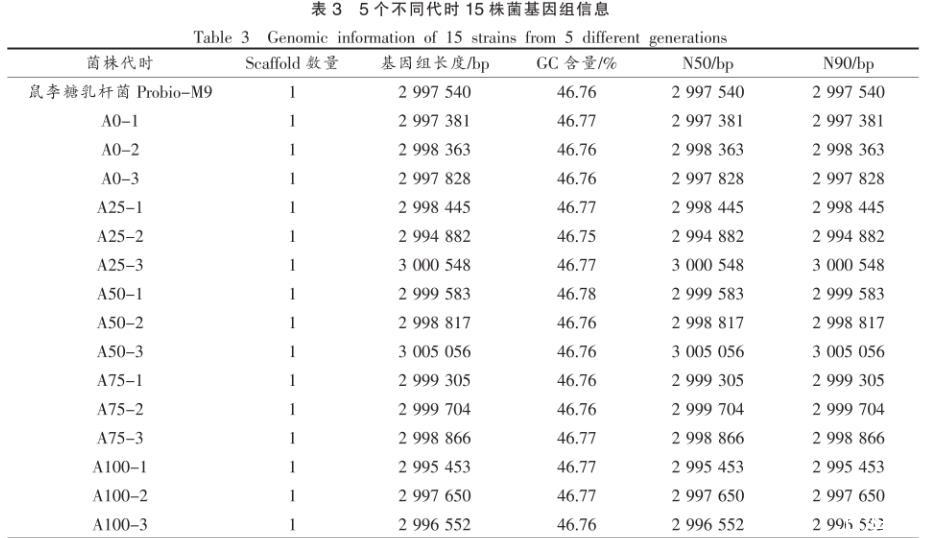
表3 5个不同代时15株菌基因组信息
2.2.2系统发育分析
本研究中利用系统发育树来描述传代过程中菌株的进化关系,进而判断菌株在连续培养过程中的稳定性。以鼠李糖乳杆菌CNCM-I-3698作为外群,使无根树转为有根树,鼠李糖乳杆菌Probio-M9原始基因组为参考基因组,使用Soapsnp方法得到的15个菌株的SNP位点连成序列,通过邻接法(NJ法)构建系统发育树。
如图4所示,系统发育树共分为两个分支,5个代时的15株菌聚在同一系统发育分支,亲缘关系极近,且明显与外群基因组处于不同分支。

图4基于SNP位点构建系统发育树
2.2.3平均核苷酸一致性分析
平均核苷酸一致性(ANI)是在核苷酸的水平上,用以比较两个基因组之间亲缘关系的常用指标,其特点是对于近缘种群的区分度更高。在比较基因组学分析中,常用ANI评估基因组间的多态性和相似性,当ANI大于95%时,即表示为同一个种。通过计算鼠李糖乳杆菌Probio-M9原始基因组和15株不同代时菌株基因组间的ANI值,并绘制成热图。
由图5可知,鼠李糖乳杆菌Probio-M9原始基因组和15株不同代时菌株基因组,两两菌株间的ANI值均在99.00%,菌株之间表现出极高的相似性,且高于种间鉴定标准,可准确鉴定为同一亚种,表明连续培养100代过程中菌株均属同一亚种即鼠李糖乳杆菌Probio-M9。
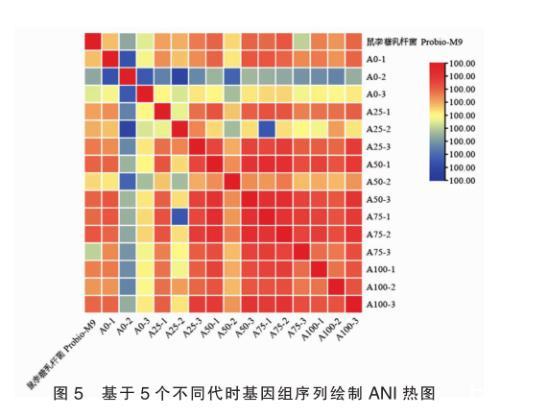
图5基于5个不同代时基因组序列绘制ANI热图
2.2.4共线性分析
共线性可用来描述同一染色体上基因的位置关系,对不同的基因组间通过基因排列顺序对其共同祖先进行探索,基因组相关性越高则其中同源序列一致性越高。本研究选取鼠李糖乳杆菌Probio-M9原始基因组序列作为参考,利用Mauve软件对15株不同代时基因组进行共线性分析。
结果如图6所示,15株不同代时培养的菌株与鼠李糖乳杆菌Probio-M9原始基因组的共线性较好,具有极高的相似性,每个基因组中均未检测到基因片段的倒位。与鼠李糖乳杆菌Probio-M9比较,A50-3和A75-2有大片段缺失,约为15 kb,主要包含整合酶家族蛋白、组氨酸激酶、核糖核酸酶、假设蛋白、tuf、cobB_1和ATP结合酶蛋白等基因。其余菌株均有小片段缺失,且基因缺失位置随机。

图6基于15株不同代时序列组装结果与鼠李糖乳杆菌Probio-M9原始基因组的共线性分析
2017年,Zhang等发现一种由asp23基因编码的碱性休克蛋白参与了干酪乳杆菌Zhang对庆大霉素的适应性,从蛋白质组学对其适应性机制进行探究,通过比较含有庆大霉素的培养基中生长的asp23基因缺失突变体及其亲本菌株的蛋白质组,发现与asp23突变株相比,一些膜相关蛋白在亲本菌株中显著上调,表明asp23编码的碱性休克蛋白能够通过调节细胞膜蛋白来适应庆大霉素的环境胁迫。本研究中15株不同代时培养的菌株均有一定的基因片段缺失现象发生,因此对于本研究中缺失片段所表达的基因功能还有待进一步研究。
2.2.5 SNP突变位点以鼠李糖乳杆菌Probio-M9为参考菌株,利用Soapsnp软件对15株不同代时鼠李糖乳杆菌Probio-M9菌株进行SNP位点识别,同时使用Samtools软件对测序数据检测到的突变位点进行过滤,剔除假阳性结果,结果见表4。

表4 15株不同代时菌株提取的SNP突变位点及注释信息
以鼠李糖乳杆菌Probio-M9原始菌株基因组为参照,鼠李糖乳杆菌Probio-M9在连续培养100代时的不同代时菌株中共发现16个SNP位点,SNP位点分布及功能如表4所示,包括6个同义突变和4个非同义突变,6个位于非编码区。非同义突变主要位于编码酪氨酸受体激酶YwqD(Tyrosine-protein kinase)和假设蛋白(Hypothetical protein)的基因上。根据突变位点对其稳定性进行分析发现,连续培养100代过程中高频突变数量较少,检测到2个SNP,且首次发生在第0代基因组与参考基因组之间,一旦建立将稳定遗传。菌株低频突变检测到的个数均小于21。研究表明,SNP个数小于21时,可将其鉴定为同一物种。不同代时15株菌所识别到的SNP位点均小于21个,表明15株不同代时菌株可鉴定为同一物种。
5个代时菌株SNP位点随机产生,从0代到25代、25代到50代、50代到75代、75代到100代的菌株中,均未发现稳定存在的突变位点。表明鼠李糖乳杆菌Probio-M9在连续传代100代时过程中,遗传稳定性良好。
2.2.6碳水化合物活性酶注释
碳水化合物活性酶数据库(CAZy)是物质对碳水化合物及其衍生物的合成及分解的酶类数据资源库。本研究通过该数据库对鼠李糖乳杆菌Probio-M9碳水化合物活性酶功能基因进行分析,在基因组水平观察鼠李糖乳杆菌Probio-M9菌株在连续培养100代时对碳水化合物利用能力的稳定程度。
如图7所示,从15株不同代时基因组中共注释到6大类碳水化合物活性酶的68个小类基因家族,主要以GHs(糖苷水解酶)和GTs(糖苷转移酶)酶类的含量最为丰富。大部分基因家族在不同代时无显著差异,虽然极个别菌株注释到的酶类存在不一致情况,但不会稳定遗传,且总体碳水化合物利用情况相同,遗传稳定性较好,与表型中不同代时碳水化合物利用能力相同的结果相吻合。
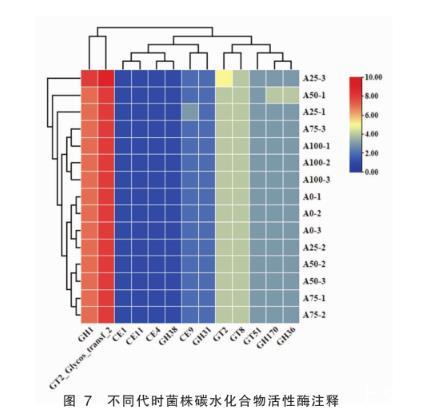
图7不同代时菌株碳水化合物活性酶注释
3、结论
遗传稳定性即子代产生的性状与亲本拥有相同的基因型。在生物进化过程中,遗传变异会经常出现,如不严格进行人工选择,就会导致菌种的衰退,从而导致菌种在生产应用过程中出现低产、不稳定现象。研究表明,菌株在培养及使用过程中,因保藏方法不同、培养基选择、传代代次和人工操作等原因会导致菌种出现衰退现象。早在1983年,Clements等发现不同批次用相同方法制备的乳杆菌在治疗腹泻的效果上存在显著的差异。引发了人们对于遗传稳定性的关注。
近年来,益生菌愈加的受大众欢迎,而在选择益生菌时,不应该仅仅考虑菌株的功能,更要参考菌株的遗传稳定性和生存能力等,这些性能关系到益生菌在肠道中的生存和定殖以及在生产应用中其益生特性的发挥。鼠李糖乳杆菌Probio-M9是2017年从中国多地的健康母乳中分离出的一株益生菌,多项研究证实,鼠李糖乳杆菌Probio-M9具有耐酸、耐胆盐特性,其显著的益生特性体现在能够活着进入人体肠道,对肠道菌群起到调节作用并增强机体免疫力,其在食品工业中广泛应用,而目前所做研究更多的停留在生理生化水平,对菌株的遗传稳定性研究少之又少。
本研究中,对鼠李糖乳杆菌Probio-M9在MRS培养基(液体培养基)中连续培养5个代时(0,25,50,75,100代)过程中的表型特征及基因组稳定性进行分析。采用全基因组及重测序方法对不同代时15株菌进行测序,基因组大小、GC含量表明15株连续培养的菌株与鼠李糖乳杆菌Probio-M9原始菌株基因组均无明显差异,系统发育关系表明15株菌与原始基因组具有较近的亲缘性。在识别到的SNP位点来看,每两个传代代时间均未发现稳定遗传的突变位点,且菌株共线性良好。同时,从基因组水平编码基因的突变水平证实了鼠李糖乳杆菌Probio-M9在连续培养100代过程中具有稳定表型特征及代谢特性。
综上所述,本试验结果表明鼠李糖乳杆菌Probio-M9在连续传代过程中具有良好的遗传稳定性,可为其后期开发应用提供理论参考。
相关新闻推荐
2、不同酵母的生长曲线、发酵力、耐冻性能测定及发酵面团感官评定(一)
3、Bioscreen C实时监测变形链球菌生物膜形成氧依赖性调控机制(一)