2.7.时序寿命分析
对于稳定期培养物,将细胞在选择性SD培养基中于30°C、220 rpm振荡培养,直至细胞密度OD600约为5。将这些稳定期细胞洗涤两次并重悬于无菌蒸馏水中,OD600为1。将细胞在37°C下振荡培养,在此温度下细胞死亡率加快。每天对培养物取样。使用碘化丙啶(PI)荧光测定法进行细胞活力分析。
2.8.碘化丙啶染色
将酵母细胞悬浮在PBS pH 7.2中,浓度约为0.2 x 10^6个细胞/毫升,然后将1μl PI储备溶液(1 mg/ml)加入0.5 ml酵母细胞悬浮液中。样品在5-10分钟内在流式细胞仪上运行,该仪器能够测量红色荧光(带通滤波器>650)。不包括洗涤步骤。
2.9.流式细胞术
在流式细胞仪上进行FACS。使用15 mW氩离子激光器,在488 nm激发波长下分析PI染色。通过675带通滤波器,通过FL3检测器测量PI作为红色信号。每个样品检测至少10,000个事件。使用CELLQuest软件进行多变量数据分析。
3.结果
3.1.人铁蛋白L改善了Dyfh1突变体的生长速率
由于FL在哺乳动物系统中具有抗氧化和铁解毒功能,我们询问FL在酵母中是否具有类似功能。酵母缺乏铁蛋白基因,并且有研究表明酵母蛋白YFH1p(人frataxin的同源物)在铁储存中发挥作用。因此,我们将人铁蛋白L链转化到酵母frataxin敲除菌株Δyfh1中并进行互补实验。通过PCR生成长度完整的人铁蛋白L(FL)基因,克隆到酵母表达载体pSCGAL10-SN(附加体质粒)中,然后转化到酵母Δyfh1菌株FKEN01501A(AL)中。作为对照,将空载体转化到Δyfh1或其亲本WT菌株中。Northern印迹分析显示,在诱导型SG培养基中FL亚基的mRNA水平很高,而在非诱导型SD和SR培养基中表达较弱(图1)。
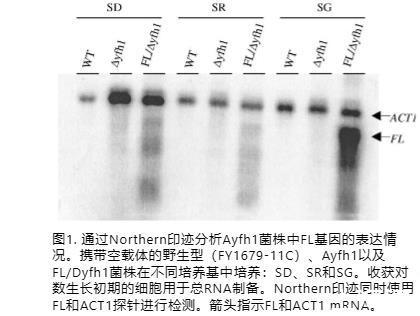
在非诱导型SD和SR培养基中观察到的低水平FL mRNA可能是由于GAL10启动子的泄漏所致,已知高拷贝数载体会发生这种情况。注意到Δyfh1菌株在SD培养基中生长良好,在SR培养基中生长缓慢,在SG培养基中生长不良(数据未显示),我们在大多数实验中使用SR培养基。通过Bioscreen测量的FL/Δyfh1和Δyfh1代表性转化子的生长速率如图2A所示。有趣的是,人FL基因的表达改善了Δyfh1突变体的生长,但并未完全恢复到WT水平。
3.2.在Δyfh1突变体中表达铁蛋白L可解毒过量的铁
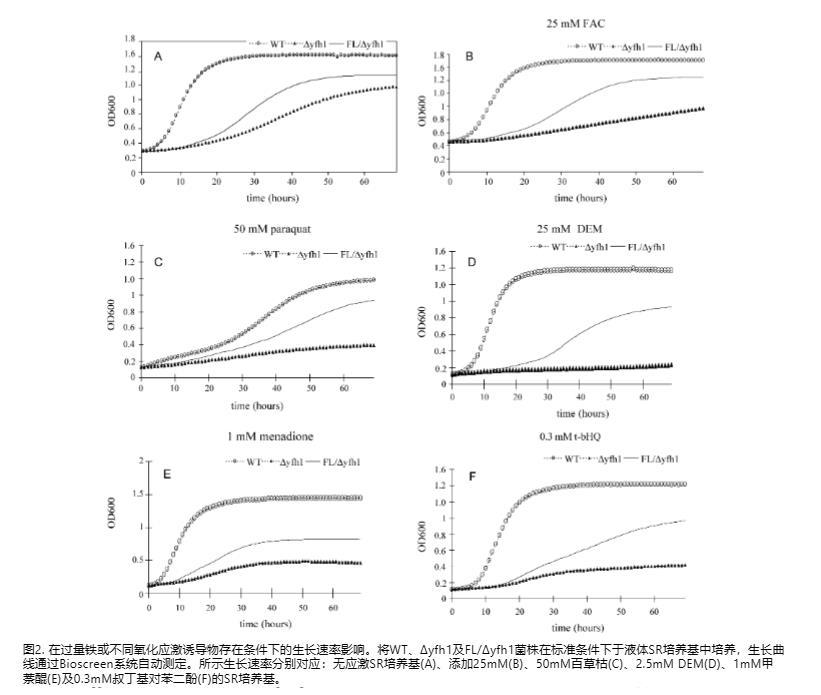
为了研究是否是FL的铁结合活性负责挽救frataxin缺陷菌株的生长缺陷,我们测量了酵母在含有过量铁的培养基中的生长速率。正如预期的那样,FL转化子显示出对过量铁的显著耐受性,而frataxin缺失突变体无法解毒铁过量并表现出生长速率降低(图2B)。FL逆转外源铁生长抑制效应的能力表明,Δyfh1中生长速率降低是由过量游离铁介导的,这可能导致ROS水平升高。
3.3.铁蛋白L保护Δyfh1菌株免受氧化剂诱导的细胞死亡
接下来我们检查FL是否也能提高Δyfh1背景菌株耐受氧化剂的能力。在存在50 mM百草枯、2.5 mM马来酸二乙酯(DEM)、1 mM甲萘醌和300μM叔丁基氢醌(t-bHQ)的情况下测量细胞生长。如图2C-F所示,Δyfh1菌株对各种氧化剂过敏。然而,FL转化子显示出对氧化应激的更大耐受性和改善的生长,尽管后者未达到野生型的水平。
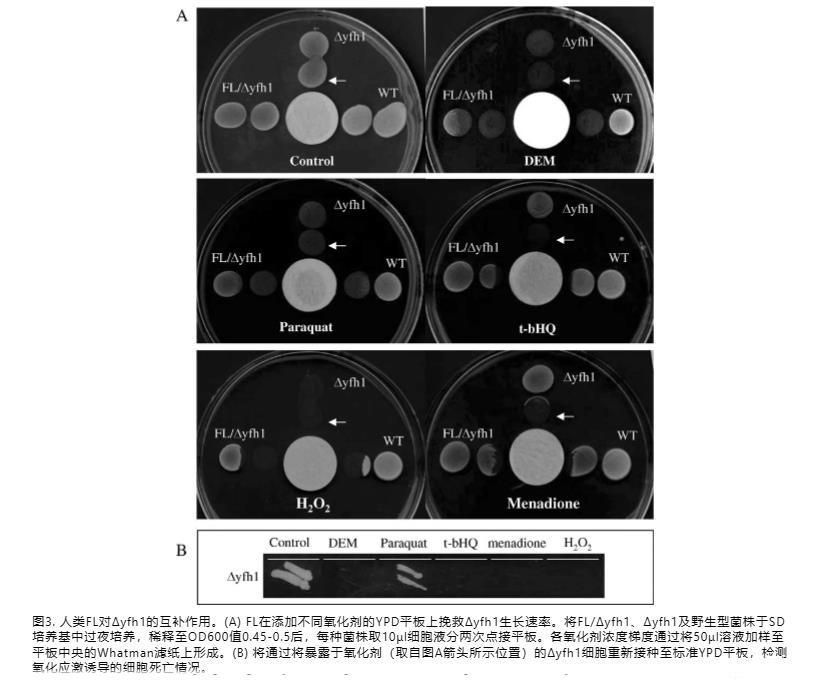
为了验证FL转化恢复生长是由于保护细胞免受氧化剂诱导的死亡,在含有氧化剂浓度梯度的固体YPD平板上分析细胞生长。通过将50μl不同的氧化剂(97%DEM、1 M百草枯、1 M甲萘醌、2M t-bHQ溶于DMSO,或3%H2O2)滴在含有YPD培养基的培养皿中央的Whatman纸上形成梯度。铁蛋白转化子比未转化的frataxin突变体表现出更强的氧化应激抵抗力;铁蛋白互补改善了细胞生长,尽管未达到WT水平(图3A)。为了区分氧化应激诱导的frataxin缺陷细胞生长停滞是由于细胞死亡还是生长抑制,我们将生长停滞的Δyfh1细胞铺板在没有应激诱导剂的YPD培养基上。3天后检查它们。先前用DEM、甲萘醌、t-bHQ或H2O2处理过的培养物中的细胞没有生长,证明这些氧化剂导致了细胞死亡(图3B)。然而,先前用百草枯处理过的培养物中的细胞生长良好,表明百草枯在不杀死细胞的情况下停止了生长(图3B)。
3.4.FL保护蛋白质免受损伤
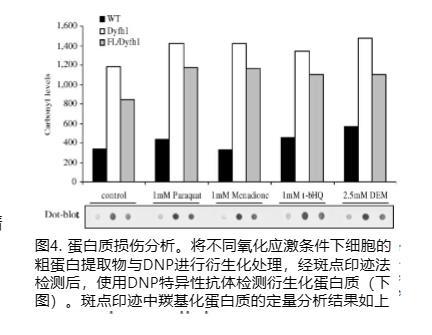
众所周知,大多数氧化剂会引起细胞蛋白质的氧化修饰。因此,我们使用点印迹分析比较了Δyfh1、WT和FL/Δyfh1中的蛋白质羰基化。图4描述了一个代表性的点印迹模式示例,该模式使用抗DNP抗体检测羰基化蛋白质。在标准或氧化应激生长条件下,缺陷型frataxin菌株显示羰基化蛋白质总量比其WT显著增加2.6-4.3倍。有趣的是,在相同的生长条件下,FL明显降低了Δyfh1菌株中羰基化蛋白质的总量,降低了17-28%。
3.5.FL部分挽救了Δyfh1突变体的寿命
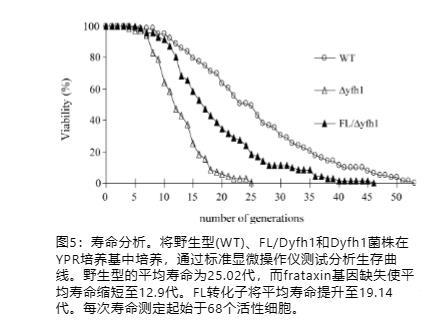
一个普遍的观察结果是氧化损伤导致衰老。酵母衰老的经典研究基于复制寿命的测量,通常通过计算母细胞在衰老前经历的分裂次数来衡量。由于frataxin突变酵母积累氧化应激,因此我们推断frataxin失活可能降低酵母的寿命,如果是这样,人铁蛋白L是否可以修复这种缺陷。为了测试这种可能性,我们在Δyfh1、其亲本菌株WT和铁蛋白转化子FL/Δyfh1中进行了寿命分析。在固体YPR培养基上进行通过显微操作的复制寿命测定,因为Δyfh1菌株在合成固体培养基上几乎不生长(数据未显示)。如图5所示,Δyfh1的复制寿命平均比WT短49%,表明frataxin确实参与酵母的寿命。有趣的是,在Δyfh1背景中转化人铁蛋白L可以挽救细胞免于死亡,并且FL/Δyfh1的寿命达到了其WT值的76.5%。
相关新闻推荐
1、水稻黄单胞菌噬菌体最佳MOI (A)、一步生长曲线、耐受性测定及基因组分析(二)