NapA是S.oneidensis硝酸盐呼吸所必需的,而NapB不是。最近有报道称napA基因是S.oneidensis将硝酸盐还原为亚硝酸盐所必需的。在本研究中,我们首先检验了napB基因是否为硝酸盐还原所必需。我们的结果表明,napB::loxP菌株能够在硝酸盐上生长(图2a),表明该蛋白对于此生物学过程并非不可或缺。鉴于该蛋白长期以来一直被认为是Nap系统所必需的,这一结果令人惊讶。有趣的是,尽管两种菌株的最大生长速率没有显著差异,但△napB菌株比野生型更早达到最大细胞密度(图2a)。为了确认这一点,对两种菌株的培养物进行系列稀释并铺在LB琼脂上进行菌落计数(图3)。与野生型相比,△napB菌株的培养物在接种后约7小时(从5小时到12小时)开始变得更多。最终,野生型达到了相同的细胞密度水平。当直接提供亚硝酸盐作为唯一电子受体时,△napA和△napB菌株表现出与野生型相似的生长曲线(图2b,数据未显示),表明NapA和NapB都不是亚硝酸盐还原所必需的。
图3:在厌氧条件下以硝酸盐为底物生长的野生型和AnapB菌株的细胞密度。每个时间点分别取各菌株培养物,进行连续稀释后涂布于LB琼脂平板进行菌落计数。相对细胞密度通过计算AnapB菌落数与野生型菌落数的比值得出。
来自△napB菌株的意外结果提出了一个可能性,即S.oneidensis可能拥有另一种硝酸盐还原途径,该途径仅在NapB缺失时才有功能。为了验证这一假设,我们构建了双突变体△napA△napB。该菌株与△napA菌株一样,无法在硝酸盐上生长或在厌氧条件下将硝酸盐还原为亚硝酸盐(图2a),表明MR-1中可能不存在替代的硝酸盐还原途径。为了证实这一观察结果,将napA基因和nap操纵子分别克隆到质粒pBBRMCS-5中进行互补。△napA△napB双突变菌株在硝酸盐上生长的能力被任一克隆的DNA片段恢复(数据未显示),表明NapA是S.oneidensis中将硝酸盐转化为亚硝酸盐的唯一硝酸盐还原酶。在以下部分中,对△napA菌株和△napA△napB双突变菌株的所有分析结果实际上相同,因此除非另有说明,使用△napA菌株代表两种突变菌株。
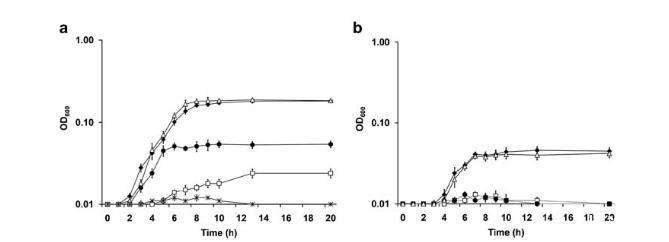
图2 野生型与突变株在厌氧条件下对硝酸盐和亚硝酸盐的生长情况。两图中均包含MR-1(◆)、ΔnapB(△)、ΔnrfA(●)和ΔcymA(□)。(a)以OD600读数表示的2mM硝酸盐生长曲线。除共有菌株外,另展示ΔnapA(*)。与ΔnapA类似,ΔnapAΔnapB无法生长(未显示)。(b)以OD600读数表示的2mM亚硝酸盐生长曲线。本图中ΔnapA=ΔnapAΔnapB=ΔnapB,为清晰起见省略ΔnapA和ΔnapAΔnapB。实验重复三次,误差棒表示均值标准差。
NrfA是S.oneidensis将亚硝酸盐还原为铵所必需的。尽管很清楚NAP系统执行硝酸盐还原为亚硝酸盐,但S.oneidensis的非典型NRF系统是否有功能仍未经过实验确定。为此,构建了△nrfA菌株。对该菌株的生理表征表明,与野生型相比,nrfA的突变导致其在硝酸盐上的生长存在严重缺陷,并且完全无法在亚硝酸盐上生长(图2b)。对于互补,质粒上的nrfA基因恢复了△nrfA菌株在亚硝酸盐上生长的能力(数据未显示)。这些结果表明NrfA是亚硝酸盐还原所必需的。有趣的是,与在两者上生长相比,S.oneidensis在单独硝酸盐(由△nrfA菌株在硝酸盐上显示)或单独亚硝酸盐(由MR-1在亚硝酸盐上显示)上的生长严重受损,表明硝酸盐还原为铵的两个步骤都有助于为代谢能量产生质子动力,从而导致更高的生物量(由更高的OD600读数指示)。
CymA替代了缺失的NapC和NrfH。Shewanella oneidensis基因组缺乏编码类似于NapC和NrfH的蛋白质的基因,这些蛋白质专门将电子传递给末端还原酶NapA和NrfA。基于迄今为止已知的NapC和NrfH都是膜结合c型细胞色素蛋白这一事实,最有可能缺失的蛋白是同一类型。早些时候,一种20.8 kDa的c型细胞色素CymA被提出是该蛋白,扮演这两个角色。然而,鉴于S.oneidensis含有40多种c型细胞色素,其中9种是细胞质膜结合的,假设没有其他c型细胞色素可能在功能上替代NapC或NrfH还为时过早。为了深入了解其他候选基因和/或验证CymA在此过程中的作用,我们重新检查了在硝酸盐与富马酸盐上生长的MR-1的转录谱(表2)。
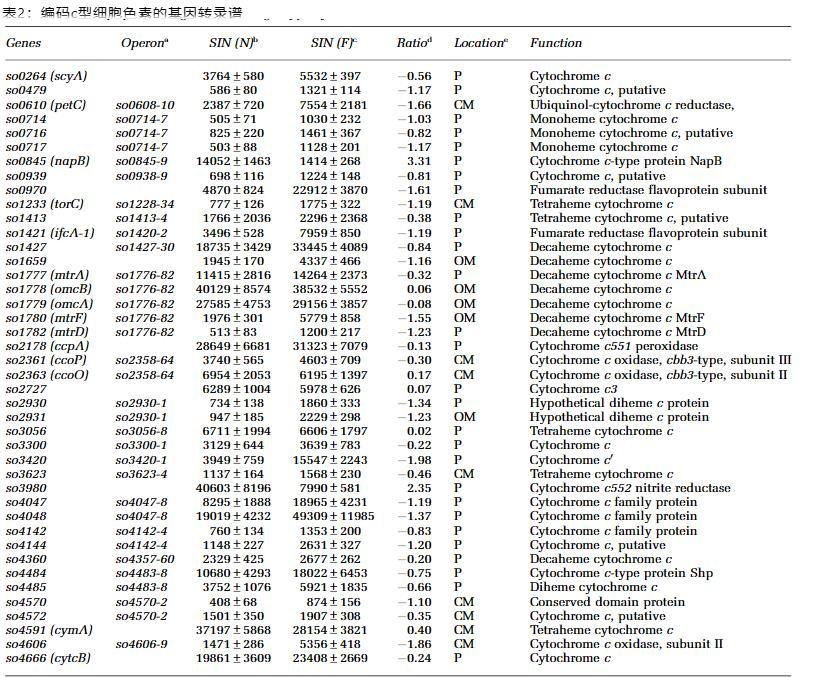
有趣的是,没有一个编码细胞质膜结合蛋白的基因被硝酸盐相对于富马酸盐显著诱导。然而,当使用硝酸盐(31797/6520,cymA信号强度/平均信号强度)或富马酸盐(28154/5521)作为电子受体时,cymA的转录水平约为所有c型细胞色素基因平均水平的五倍。先前当使用包括氧气在内的多种化学物质作为唯一电子受体时,已观察到cymA的高水平组成型表达,这被认为与其在电子传递中的关键作用有关。
尽管CymA是通过突变分析在S.oneidensis中研究最为深入的蛋白质之一,但鉴于其在厌氧呼吸中的特殊重要性,其生理功能仍需重新评估。研究人员构建了cymA基因框内缺失突变株,该DcymA菌株无法在富马酸盐、二甲基亚砜、三价铁(Fe(III))、四价锰(Mn(IV))或亚硝酸盐上生长(图2b),这与早期研究结果一致。然而,虽然DcymA菌株在硝酸盐上的生长能力较野生型存在严重缺陷,但与DnapA菌株相比,其OD600读数仍显示出微小但显著的提升(图2a)。综合来看,CymA极可能是通过NapB或直接向NrfA传递电子的主要且唯一蛋白质。
NapB是CymA向NapA传递电子的首选载体,但不参与NrfA的电子传递。DnapB菌株在生长初期细胞密度显著增加的现象令人好奇。为探究DnapB菌株的代谢特征及硝酸盐还原途径的特性,我们对野生型、DnapA、DnapB、DnapADnapB、DnrfA、DcymA和DnapBDnrfA菌株的全细胞硝酸盐还原为亚硝酸盐、亚硝酸盐还原为铵的速率进行了定量计算与比较。所有速率均经样品蛋白浓度标准化处理,结果详见表3,还原动力学过程如图4所示。与先前报道一致,野生型菌株中硝酸盐还原为铵的过程实际上为两步反应,其中亚硝酸盐还原阶段需满足特定条件才会启动。
相关新闻推荐
1、嗜碱盐单胞菌菌株生理生化与生长特性、最优发酵条件——结果与分析