使用ChIP筛选BfmA调控的下游基因。BfmA-BfmK TCS的调节子以前从未被研究过。我们使用STRING数据库预测BfmA和BfmK可能的功能伙伴。根据基因组邻域、共现或不同物种中的基因融合事件的信息判断,BfmK和BfmA各自可能与10个蛋白质相互作用,这些蛋白质的得分很高(得分>0.850),其中三个是其他HK(Smlt0977[PhoR],RpfC[Smlt2234],和SmeS[Smlt4477])。这些蛋白质中的六对,即PhoB-PhoR,Smlt1218-Smlt1219,Smlt1540-Smlt1541,Smlt3944-Smlt3949,SmeS-SmeR,和BfmA-BfmK,本身在基因组中成簇存在,表明与TCS BfmA-BfmK有很强的功能相关性。
由于BfmA是一个典型的OmpR家族RR,其C端具有一个转录因子(TF)区域(151至228个氨基酸)作为输出结构域,我们提出这些相关蛋白编码基因中的一些在转录水平上受BfmA直接调控。因此,通过DOOR数据库预测了这些相互作用蛋白编码基因的15个操纵子结构,并根据这些操纵子的预测启动子区域设计了PCR引物。然后使用ChIP结合半定量PCR来筛选可能与TF BfmA结合的DNA。
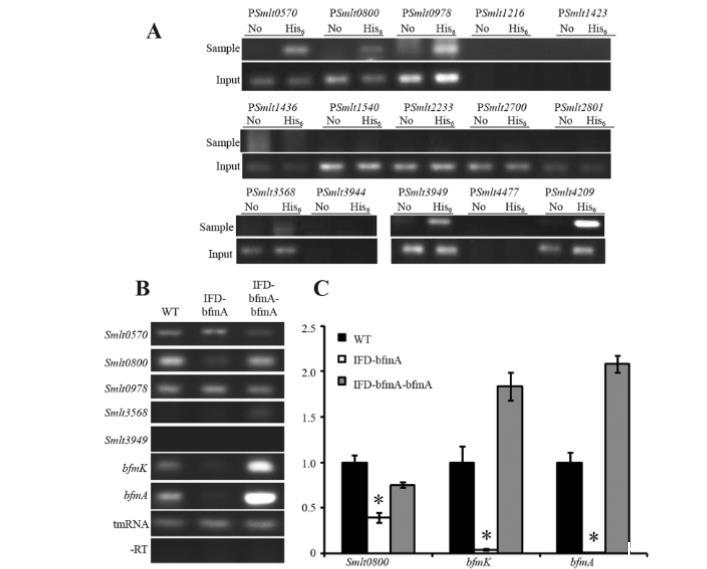
图5 BfmA调控其自身操纵子及Smlt0800的转录。(A) 通过染色质免疫沉淀(ChIP)技术筛选BfmA结合序列。细菌菌株在NYG培养基中培养至OD600值为0.4,采用ChIP收集BfmA结合的双链DNA。使用半定量PCR对与BfmA共沉淀的DNA量进行定量。No:未标记菌株(IFD-bfmA-bfmA)的ChIP DNA样本;His6:His6标记菌株(IFD-bfmA-bfmA*)的ChIP DNA样本;Sample:使用ChIP DNA进行PCR扩增;Input:使用ChIP前从细胞裂解液提取的细菌DNA进行PCR扩增(作为上样对照)。(B) 候选基因转录水平的半定量RT-PCR分析。cDNA使用嗜麦芽窄食单胞菌ATCC 13637的总RNA反转录合成。-RT表示阴性对照(cDNA合成过程中未加入反转录酶)。所有测试菌株均携带pBBRMCS2载体以确保可比性。实验独立重复三次。(C) 通过实时定量PCR检测BfmA调控基因的转录水平。以tmRNA扩增的cDNA作为内参。每个检测均采用生物学重复和技术重复三次。图示为代表性实验结果。竖线表示标准差。*表示经Student t检验计算的统计学显著差异(P<0.05)。
为进行此实验,我们构建了另一个bfmA互补菌株,类似于IFD-bfmA-bfmA菌株,但在bfmA序列的终止密码子5'区添加了一个His6表位标签编码序列。将该DNA插入片段克隆到pBBRMCS2载体中,用于互补bfmA突变体(菌株IFD-bfmA-bfmA)。使用针对BfmA-His6的单克隆抗体,基于该重组菌株(IFD-bfmA-bfmA)进行ChIP。如图5A所示,即使使用输入DNA样本作为DNA上样对照,PSmlt1216、PSmlt1423、PSmlt3944和PSmlt4477的启动子区域也未能通过PCR扩增,从而表明这些顺式调控序列中存在大量的遗传多态性。此外,PSmlt1436、PSmlt1540、PSmlt2233、PSmlt2700和PSmlt2801的PCR产物在样品中缺失,表明这些启动子不与BfmA结合。然后将上述九个启动子区域从后续分析中排除。然而,与使用来自缺乏His6标签的IFD-bfmA-bfmA对照菌株的共免疫沉淀DNA进行PCR扩增的背景对照相比,来自IFD-bfmA-bfmA*ChIP样品的PSmlt0570、PSmlt0800、PSmlt0978、PSmlt3568、PSmlt3949和PSmlt4209 PCR产物的量显著增加(图5A),表明BfmA-His6在体内直接结合这些启动子区域,并且相应的基因或操纵子受BfmA控制。
bfmA正向调控其自身操纵子和Smlt0800的转录
为了评估BfmA对其候选下游基因的调控效果,选择了其启动子区域可被BfmA结合的六个操纵子,并通过半定量RT-PCR比较了野生型菌株和IFD-bfmA突变体中mRNA的量。如图5B所示,未观察到代表性基因Smlt3949和Smlt3568的阳性扩增条带,表明在本研究使用的生长条件下它们的转录水平太低而无法检测到。Smlt0570和Smlt0978的mRNA量在测试的菌株中保持稳定(图5B),实时定量PCR也显示它们的mRNA水平在野生型菌株和bfmA突变体之间相似(见补充材料中的图S4)。这些结果表明,尽管存在TF-启动子结合事件,这两个基因的转录与BfmA的控制无关。Smlt0800和bfmK的转录水平在IFD-bfmA突变体中显著降低,而bfmA的遗传互补将其转录水平恢复至接近或高于野生型水平,表明BfmA作为正调控因子调节它们的表达。
为验证上述结果,采用实时定量PCR比较Smlt0800和bfmK的mRNA量。获得的结果与半定量RT-PCR分析的结果非常一致(图5C);bfmA的框内缺失导致Smlt0800和bfmA的mRNA量分别减少60%和96%,并且这些减少几乎完全被突变体中bfmA的遗传互补所抑制(图5C)。总之,结果表明bfmA正向自动调节其自身操纵子的转录。此外,BfmA也是一个正调控因子,控制Smlt0800的转录,该基因编码一个参与酰基-CoA水解的酰基-CoA硫酯酶(命名为AcoT)。
相关新闻推荐
1、斜带石斑鱼哈维氏弧菌分离纯化、生长曲线绘制及耐药性研究(二)