与纤毛虫相反,阿米巴被认为是专门的表面取食者。在我们的实验中,表面取食效应在生物膜生物量上可见。然而,这种减少在我们实验的最后三周最为明显(图2B)。值得注意的是,阿米巴也能够显著减少自由水中的细菌生物量(图2A和表1)。一种解释是阿米巴通过胞饮作用消耗了悬浮细菌。这种与纤毛虫的生态位重叠也与观察到的纤毛虫种群动态一致:纤毛虫种群大小在单一敌害处理中平均高于与噬菌体和阿米巴混合的处理,并且在多敌害处理中,纤毛虫密度在阿米巴种群下降后短暂增加(图4A)。阿米巴的下降是严重的,因为细胞在1周后无法用暗场显微镜检测到(图4B)。尽管暗场显微镜是在正常条件下观察阿米巴细胞的好工具,但在饥饿条件下计数阿米巴细胞可能不够充分。因此,阿米巴密度可能被低估了,特别是在食物竞争激烈的条件下,即阿米巴种群可能以非典型形式在多敌害处理中持续存在于整个实验期间。阿米巴的急剧下降也表明纤毛虫和阿米巴之间的竞争是不对称的,有利于纤毛虫,已知纤毛虫在类似的微观世界条件下是有效的细菌捕食者。
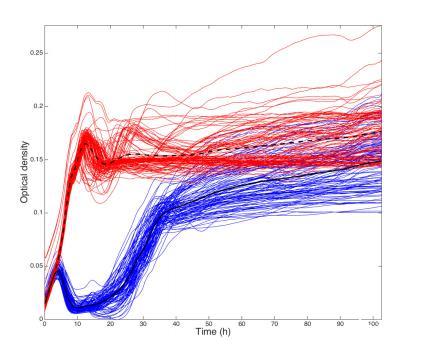
与捕食者相反,裂解性噬菌体对开放水中细菌密度的长期影响最小,并且对附着生物膜的量没有影响。然而,当观察Semad11和Db11之间的短期动态时,噬菌体被证明在杀死其宿主方面极其有效。我们独立的短期实验证明了细菌种群在12小时内初始下降了93%。高死亡风险可能对宿主防御产生强大的选择压力,导致细菌与噬菌体卷入紧密的共同进化军备竞赛动态,其中特定宿主和寄生虫基因型的适合度取决于时间。与我们实验相对应的动态也被观察到;细菌丰度的初始快速下降只是暂时的,因为具有噬菌体抗性的细菌能够在100小时内将种群恢复到接近之前的水平(图3)。因此,观察到的噬菌体抗性的出现似乎有效地削弱了寄生噬菌体与其宿主细菌之间的营养联系。
这一发现也展示了噬菌体疗法面临的主要挑战之一——防御的快速进化。然而,结果也表明,尽管噬菌体抗性细菌很快出现,但细菌种群从未能够达到单独培养细菌的对照组水平(图2A)。这可能是因为新的噬菌体基因型的出现限制了总细菌种群,推测是进化军备竞赛的结果。或者,噬菌体抗性可能是表型的(例如,细胞表面噬菌体受体的掩蔽),,导致与生长能力的权衡。尽管已经表明,使用噬菌体混合物而不是单一噬菌体可以延迟噬菌体抗性细菌变异的出现,这为我们多敌害实验方法带来了有趣的未来前景。
已知噬菌体抗性的成本取决于几个因素,例如噬菌体的遗传背景、感染期间噬菌体菌株的数量以及环境条件。例如,在土壤和池塘水等低营养环境中,噬菌体抗性的成本可能很高。因此,限制细菌生长的适合度成本可以解释噬菌体在我们实验中对自由水相生物量产生的微小但持续的长期负面影响。
阿米巴和纤毛虫的种群大小在实验期间缓慢下降。在类似捕食者-猎物系统的先前研究中发现了类似的结果,这种下降可能归因于猎物防御的进化增加,而几乎没有证据表明捕食者发生了共同进化。或者,由于实验设置中未知的因素,培养条件可能已经恶化,导致阿米巴和纤毛虫种群下降。然而,与噬菌体不同,潜在的防御进化与捕食者控制细菌种群的能力没有明显关联。在我们的实验中,我们还发现多敌害系统中纤毛虫的种群大小波动更大,这表明在自然界中,由于不同细菌消费者的相互作用,情况可能更为复杂(图4A)。在我们的案例中,波动增加可能是由于阿米巴种群规模的下降,这是由于纤毛虫的激烈竞争。此外,已经证明,在存在捕食者的情况下,宿主-寄生虫的共同进化与没有捕食者时不同,因此可能也存在其他生态进化原因导致观察到的波动。
总之,我们的结果表明,对细菌猎物的下行效应在很大程度上取决于敌害的取食模式。纤毛虫是非常有效的开放水体捕食者,而阿米巴能够减少细菌生物膜和开放水体生物量。这可能导致资源利用的部分和不对称重叠,并可能导致阿米巴的减少和观察到的纤毛虫种群动态的短期效应。我们还发现,虽然噬菌体在短期是超级有效的寄生虫,但噬菌体的长期影响可以忽略不计。这一结果源于噬菌体抗性的极其快速的出现以及噬菌体抵消它的能力较弱。我们的结果强调,尽管来自细菌及其捕食者或寄生虫的单敌害实验数据很有价值,但由于敌害之间的相互作用,它们可能无法反映多敌害食物网中发生的过程。此外,敌害和宿主之间快速的进化动态可以极大地影响捕食性原生动物与寄生噬菌体作为细菌丰度和空间分布调节者的相对重要性。
从细菌病原体的生物防治角度来看,我们的结果强调,单一的裂解性噬菌体在短期内可能有效,而纤毛虫和阿米巴对细菌具有更持久的影响。尽管如此,我们建议噬菌体和原生动物一起可以为环境生长细菌的可持续和安全抗菌疗法提供一种新选择。此类细菌的一个例子是在水产养殖中具有主要意义的鱼类病原体柱状黄杆菌,它是柱状病的致病因子,每年给鱼类养殖造成重大的经济损失。