产肺炎碳青霉烯酶的铜绿假单胞菌(KPC-PA)分离株在中国迅速扩增,尤其是高危克隆ST463。旨在探索推动ST463克隆成功的KPC相关质粒的进化。本研究对2011—2020年收集的1258株临床铜绿假单胞菌菌株进行全基因组测序,识别出106株ST463-PA菌株,其中KPC的检出率为90.6%。研究发现在早期(2011—2012年),ST463-PA获得了编码KPC的II型(pT2-KPC)或I型质粒(pT1-KPC)以克服碳青霉烯类抗生素的压力。2012—2017年间,由于pT1-KPC质粒具有较低的适应性成本和IS26驱动的blaKPC扩增能力,其成为主导质粒。到2017—2020年,pT1-KPC质粒发生大片段缺失,形成了pT1del-KPC质粒。该质粒具有更低的适应性成本、增强的blaKPC-2基因稳定性和更大的拷贝数灵活性,同时保持了水平传播能力。因此pT1del-KPC质粒最终取得成功,使ST463成为中国的主要ST。研究还评估了不同质粒模式在ST463遗传背景下的适应性成本,并发现携带pT1del-KPC质粒的菌株在无抗生素或高浓度美罗培南条件下比携带pT1-KPC质粒的菌株具有更高的适应性。在低浓度美罗培南压力下,pT1del-KPC质粒通过增加质粒拷贝数来放大blaKPC-2基因,而pT1-KPC质粒则通过IS26介导的blaKPC-2基因扩增来响应。这些发现强调了进化压力推动ST463优势的形成,并强调了需要有针对性的策略来控制其传播和抗生素抗性的发展。
Bioscreen全自动微生物生长曲线分析仪的应用
Bioscreen全自动生长曲线分析仪主要用于获取不同抗生素浓度下不同铜绿假单胞菌分离株的生长曲线。首先将1.5μL过夜培养的菌液稀释到998.5μL含有或不含抗生素的阳离子调整Mueller–Hinton肉汤(CAMHB)中,将稀释后的样本在37°C下使用Bioscreen全自动生长曲线分析仪主连续振荡培养,并每5分钟测量一次420-580nm波长下的光密度值,持续1300分钟。每个实验设置进行了三组生物学重复,每组生物学重复又进行了三组技术重复。评估不同质粒模式下铜绿假单胞菌在不同抗生素压力下的生长适应性。
实验结果
研究结果表明,中国和美国的ST463菌株在blaKPC基因和质粒携带方面存在显著差异。在中国,几乎所有ST463菌株都携带I型和/或II型质粒(446/463),其中大部分还携带blaKPC基因(433/463)。相比之下,美国的ST463菌株中只有3株携带I型或II型质粒,且没有菌株携带blaKPC基因。这表明ST463可能起源于美国,随后传播到中国,在中国进化出KPC质粒,促使其在中国多个医院迅速传播。KPC相关质粒在ST463-PA中的多阶段进化轨迹。在第一阶段(2011—2012年),原始ST463菌株通过在原始pT1质粒中整合blaKPC-2基因(形成模式A)或获得pT2-KPC质粒(形成模式B)来克服碳青霉烯类压力。在第二阶段(2012—2017年),由于pT1-KPC质粒比pT2-KPC质粒具有更高的适应性,携带pT1-KPC质粒的菌株(模式A)在ST463 CRPA中变得普遍。在第三阶段(2017—2020年),pT1del-KPC质粒(模式C)为ST463菌株提供了比pT1-KPC质粒(模式A)更大的适应性优势,从而取代了模式A,使ST463成为CRPA中的主要ST。携带pT1del-KPC质粒的菌株在无抗生素或高浓度美罗培南条件下比携带pT1-KPC质粒的菌株具有更高的适应性。在低浓度美罗培南压力下,pT1del-KPC质粒通过增加质粒拷贝数来放大blaKPC-2基因,而pT1-KPC质粒则通过IS26介导的blaKPC-2基因扩增来响应。
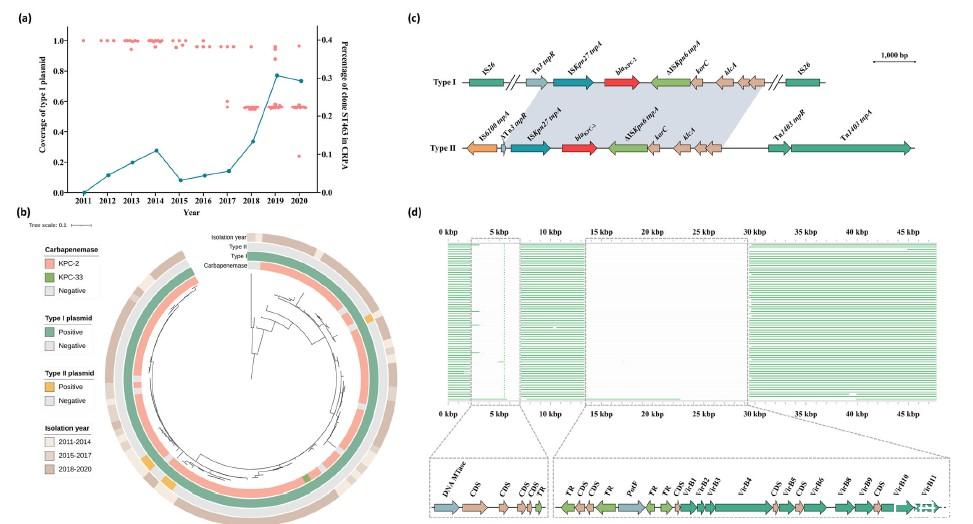
图1、本研究纳入的106株ST463-PA分离株的基因组特征。a)ST463-PA分离株中I型质粒覆盖率(针对pS11-17进行爆破)的分布(以红点表示);以及克隆ST463在CRPA分离株中的患病率(以蓝线表示)。b)ST463-PA分离株的核心基因组系统发育树。碳青霉烯酶类型、I型质粒的存在、II型质粒的存在和分离年份由内而外表示。c)比较ST463-PA分离株中I型和II型质粒的典型blaKPC-2遗传背景。灰色阴影表示相同的序列。d)来自ST463-PA分离株的质粒重叠群比对显示对pS11-17的覆盖率降低。CDS编码序列,CRPA碳青霉烯类耐药铜绿假单胞菌,TR转录调节因子。
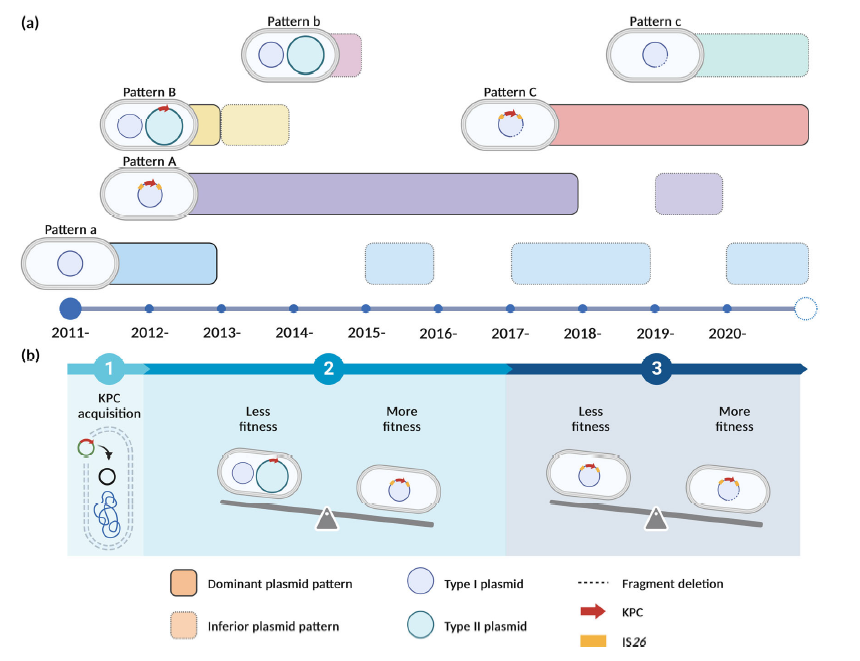
图2、ST463-PA分离株中KPC相关质粒的进化过程。a在研究中,来自ST463-PA分离株的不同KPC相关质粒模式的时间分布。每年的显性和从属质粒模式分别以实线的不透明色和虚线的透明色为标志。b关于ST463-PA分离株中KPC相关质粒多阶段进化轨迹的提案。
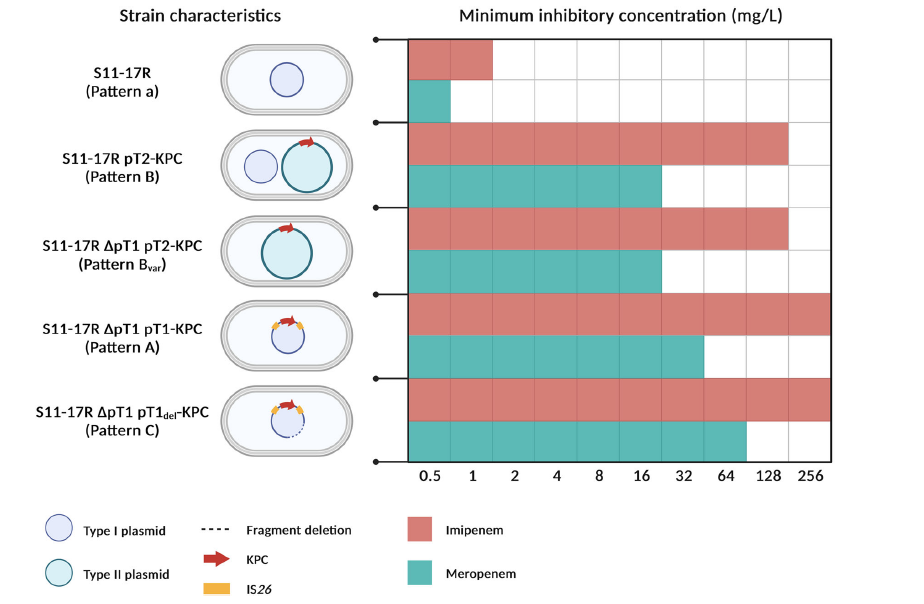
图3、本研究中使用的实验性ST463菌株的特征。实验菌株及其相应的碳青霉烯类最低抑制浓度的示意图如图所示
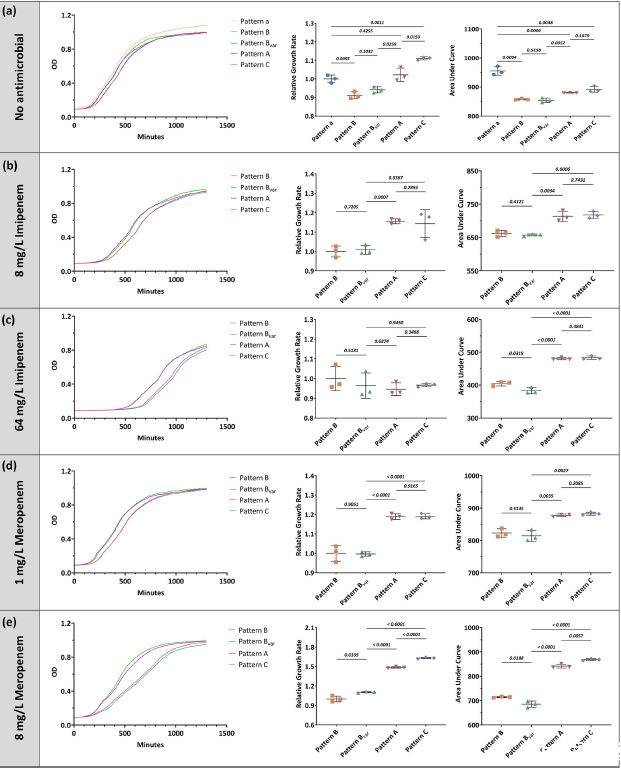
图4、实验性ST463菌株中与不同KPC相关质粒相关的适应性成本。测试了各种条件下的生长过程:a)无抗生素、b)8 mg/L亚胺培南、c)64 mg/L亚胺培南、d)1 mg/L美罗培南和e)8 mg/L美罗培南。面板中的每个数据点对应于从三个技术重复(n=3个生物独立样本)计算出的一个生物重复。
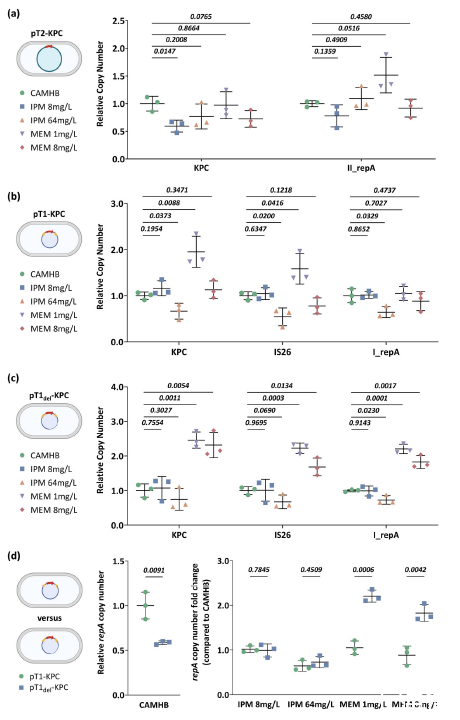
图5、不同碳青霉烯类压力下的基因拷贝数变化。a-c)pT2-KPC、pT1-KPC和pT1del-KPC质粒的基因拷贝数。d)pT1-KPC和pT1 del-KPC质粒之间的比较。面板中的每个数据点对应于从三个技术重复(n=3个生物独立样本)计算出的一个生物重复。
总结
产肺炎碳青霉烯酶的铜绿假单胞菌(KPC-PA)分离株在国内迅速扩增,尤其是高危克隆ST463。本研究旨在探索推动ST463克隆成功的KPC相关质粒的进化。对1258株临床铜绿假单胞菌菌株的全基因组测序鉴定出106株ST463-PA分离株,KPC患病率为90.6%。早期(2011-2012),ST463-PA获得了编码KPC的II型(pT2-KPC)或I型质粒(pT1-KPC)来克服碳青霉烯类应激。2012年至2017年间,pT1-KPC质粒因其较低的适应性成本和IS26驱动的blaKPC扩增能力而占主导地位。到2017-2020年,pT1-KPC中的大片段缺失形成了pT1del-KPC质粒。它赋予了更低的适应性成本、增强的blaKPC-2基因稳定性和更大的拷贝数灵活性,同时保持了水平传播能力。因此pT1 del-KPC质粒最终成功,使ST463成为中国优势ST。研究结果强调了推动ST463优势的进化压力,并强调需要有针对性的策略来控制其传播和抗生素耐药性的发展。通过Bioscreen C全自动生长曲线分析仪测量得到的生长曲线数据,研究人员能够清晰地观察到不同类型和状态的KPC质粒在铜绿假单胞菌中的生长动态。例如在没有抗生素的情况下,携带特定质粒模式的菌株可能生长缓慢,显示出较高的适应性成本。而在有抗生素压力的条件下,某些质粒模式的菌株能够更好地生长,表明这些质粒为菌株提供了对抗抗生素的适应性优势。这些发现对于理解KPC质粒在铜绿假单胞菌中的进化和传播具有重要意义,也有助于揭示质粒结构变化如何影响菌株的生长和抗生素抗性。
相关新闻推荐
2、益生菌靶向肠道微生物群,改善血脑屏障及其相关疾病的研究(三)