摘要
通过使用特定的半固体培养基和分光光度计/比浊测量方法,结合Bioscreen C的2x100微孔板,开发了一种快速筛选抗真菌化合物和进行丝状真菌生态生理学研究的方法。培养基组成和制备、接种物大小、培养基体积以及用于测量初始萌发和生长动态的培养参数均已优化。这些方法已应用于评估18种浓度丙基丙烷硫代亚磺酸酯(PTS)在YES培养基中对黄曲霉(Aspergillus flavus)在不同环境条件下的有效性。在四种不同环境条件(水活度0.995和0.95;温度20°C和25°C)下,通过7天周期内每20分钟自动测量,计算了最小抑制浓度(MIC)、非抑制浓度(NIC)和新开发的MIC50。数据使用Lambert-Pearson模型进行建模,这是一种先前用于细菌抑制的数学建模方法。结果已与传统生长速率数据和半致死剂量(LD50)值进行比较。这种方法可能对丝状生物的生长和次级代谢产物生产的快速筛选assay产生重大影响,并在时间减少、所需培养基体积、测量以及整合和建模关键参数以比较效能方面具有主要优势。
引言
确保食品微生物质量和安全的需求促进了预测建模在量化和预测微生物行为方面的兴趣(Lahlali等人,2005)。特别令人关注的是不同环境因素和防腐剂对生长前滞后期和生长速率的影响。由于丝状真菌的生长习性,尤其是产毒菌种,进行此类实验更为困难,因此环境因素相互作用对生长和边界条件影响的建模研究较细菌受到较少关注(Garcia等人,2009)。
真菌活性(包括孢子萌发、生长和孢子形成)的量化是复杂的,因为真菌通过菌丝在基质中定殖。真菌产生菌丝生物量,其重量仅在生长早期阶段呈指数增加(Koch,1975),以及它们不是单细胞,并能在三维空间中扩散和穿透食品基质,使得生长量化和建模困难(Gibson&Hocking,1997;Dantigny等人,2005)。这些障碍导致使用菌丝延伸速率(如径向生长速率)作为最常见的方法,基于Trinci(1971)的工作。然而,这些测量并不代表真菌生长的真实三维性质。
研究环境因素(培养基、水可用性、温度)和筛选抗真菌化合物(尤其是食品腐败产毒真菌)的生态生理学研究通常使用基于琼脂的平皿系统传统测量方法(Marin等人,1995;Medina等人,2007;Judet等人,2008)。这些多因素实验通常需要长时间、大量重复平皿,并需要定期测量菌落直径。
在大多数情况下,丝状真菌的生长动力学是从菌丝延伸速率计算的。对于抗真菌效能,这些数据然后用于计算相对于对照抑制50%(LD50)和90%(LD90)生长的致死剂量(Aldred等人,2008;Kumar等人,2010)。这通常只允许测试少量浓度,并需要插值,假设响应是线性的。
基于比浊测量的分光光度法assay方法已在少量真菌生长研究中使用(Schnurer,1993;Rossi-Rodrigues等人,2009)。这些研究主要在医学领域(Van Eldere等人,1996;Meletiadis等人,2001a,2001b;Meletiadis等人,2003),并用于开发分生孢子形成真菌的体外抗真菌敏感性测试;这提供了快速结果,以光密度(O.D.)单位表示。
使用比浊测量获得的数据已被许多作者用于计算两个对食品工业和评估目的特别感兴趣的特定浓度。这涉及获取非抑制浓度(NIC)和最小抑制浓度(MIC)在特定环境条件下的信息(Lambert&Pearson,2000)。此类丝状真菌研究通常持续24至48小时(Schnurer,1993;Meletiadis等人,2001a,2001b;Rossi-Rodrigues等人,2009)。据我们所知,尚未有更长期研究的描述。问题通常是较长培养期导致生长曲线不规则,并且因为曲线下面积(AUC)是用于计算参数。这一限制使得该技术不适合丝状真菌生长的扩展生态生理学研究。
Lambert&Pearson(2000)和Lambert&Lambert(2003)的研究导致了适当的数学工具开发,用于计算单一和混合抑制剂对细菌的MIC和NIC值。Lambert-Pearson模型(LPM)(方程(1))已成功用于O.D.(Lambert&Pearson,2000;Lambert&Lambert,2003;Bidlas&Lambert,2008)和阻抗微生物测量(Chorianopoulos等人,2006)。这种数学方法使用检测时间(TTD)计算MIC(方程(2))和NIC(方程(3)),这使得计算独立于实验时间。

其中RTD是检测速率(1/TTD),x是抑制剂浓度,P0是最优RTD值,P1和P2是最大斜率浓度(在对数浓度轴上)和斜率参数,e是指数,约等于2.718。
本研究的目标是(1)开发一种自动比浊读数器方法检查黄曲霉的萌发和初始生长动态,(2)使用所提出的方法结合LPM准确定量评估抗真菌化合物在不同环境条件下对黄曲霉的MIC和NIC。
材料和方法
真菌菌株和培养基
使用一株产黄曲霉毒素的黄曲霉菌株(NRRL 3357),由美国农业部(USDA)新奥尔良的D.Bhatnagar博士提供。该菌株在麦芽提取物琼脂(20克麦芽提取物(Difco),2克蛋白胨(Difco),15克琼脂(Sigma-Aldrich,多塞特,英国))上于25°C黑暗培养7天。使用YES琼脂培养基,含酵母提取物20克/升,蔗糖150克/升和硫酸镁0.5克/升。用于制备培养基的水用甘油修改至所需水活度水平(aw;0.995和0.95)。
化学品
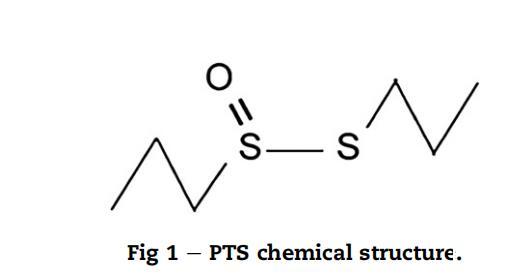
丙基丙烷硫代亚磺酸酯(PTS)(图1),一种从大蒜球茎(Allium sativum)中初始化合物分解获得的有机磺酸盐,由DMC研究中心S.L.,格拉纳达,西班牙(X.Paris博士)提供。储备溶液通过将纯PTS稀释在80%乙醇水溶液(v/v)中制备。工作标准系列在每次实验前制备,以便所有培养基接种相同量的乙醇。不含PTS的80%乙醇溶液在YES培养基中用作对照培养基。
比浊assay
浊度分析使用Bioscreen C微生物生长分析仪(Labsystems,赫尔辛基,芬兰)。非标准100孔微孔板专为此机器制造,加载先前接种黄曲霉孢子悬浮液的不同培养基。O.D.使用600纳米滤光片每20分钟记录一次,持续7天。数据使用制造商提供的软件Easy Bioscreen Experiment(EZExperiment)记录,然后导出到Microsoft®Excel®Professional 2010(14.0.4756.1000)(Microsoft Corporation,雷德蒙德,华盛顿,美国)表格进行进一步分析。实验在20°C或25°C下进行。
培养基制备和琼脂浓度优化
使用YES培养基。使用纯水,测试六种不同琼脂浓度(2-0.05%w/v)。培养基在121°C高压灭菌20分钟。之后,培养基摇动并在室温下完全冷却。培养基(300μl)使用多通道移液器倒入100孔微孔板(每种琼脂浓度十个孔)。Bioscreen C在25°C下运行45小时。
选择适当接种物大小
使用在YES琼脂培养基上生长的黄曲霉菌株(7天,25°C)制备含1.1x10^8孢子/毫升的孢子悬浮液,在0.05%Tween 80盐溶液中。制备系列稀释以获得从10^3到10^7孢子/毫升的不同孢子浓度。这些溶液用于接种半固体YES培养基(含先前部分最佳琼脂浓度),浓度范围从10^2到10^6孢子/毫升。然后培养基使用多通道移液器倒入100孔微孔板。每种处理使用十个重复。
选择适当体积
半固体YES培养基(0.125%琼脂w/v)接种黄曲霉孢子悬浮液以获得最终浓度1x10^5孢子/毫升,使用不同体积(200至400μl)加载到100孔微孔板孔中。每种条件使用十个重复。
不同O.D.值下生物量计算
获得两个含1x10^6和1x10^8孢子/毫升的孢子溶液。使用两组半固体YES培养基(40毫升)接种微孔板,每孔加入400μl一种孢子悬浮液,给出最终浓度1x10^4和1x10^6孢子/毫升。微孔板五十孔填充每种混合物,然后在25°C培养。
培养13、14、15、16和17小时后,记录O.D.,并使用无菌1毫升注射器将十孔内容物提取到预称重的塑料离心管中。菌丝和孢子通过添加3毫升无菌双蒸水洗涤四次,摇动后,以6000 rpm离心20分钟。之后,管在无菌流动柜中干燥,再次记录重量。通过减去原始管重量计算存在的生物量。
相关新闻推荐
2、生长曲线分析仪:测定不同TIR防御系统在大肠杆菌中表达时对噬菌体感染的反应
3、白色念珠菌来源的胞外囊泡EVs对粪肠球菌生长和致病性的影响(下)