研究简介
本研究揭示了假单胞菌(Pseudomonas putida S16)中尼古丁降解的未知途径。尼古丁是一种常见的有机污染物,主要来源于烟草加工废弃物。假单胞菌能够利用尼古丁作为碳源和氮源进行生长,但其降解尼古丁的分子机制尚不清楚。
本研究人员通过蛋白质组学和分子遗传学方法,全面分析了假单胞菌在尼古丁和甘油(非抑制性碳源)培养条件下的蛋白质表达差异,揭示了尼古丁降解的关键酶和基因。研究发现,假单胞菌在尼古丁培养条件下表达的蛋白质组包含1292种蛋白质,其中126种蛋白质在尼古丁和甘油培养条件下表达差异显著。这些差异表达的蛋白质主要涉及运输、解毒、氨基酸代谢等功能。研究还发现,3-琥珀酰吡啶(SP)转化为6-羟基-3-琥珀酰吡啶(HSP)的关键步骤由一种多酶反应催化,涉及钼蝶呤结合氧化酶(spmA)、钼蝶呤脱氢酶(spmB)和(2Fe-2S)结合铁氧还蛋白(spmC),并以钼蝶呤胞嘧啶二核苷酸为辅因子。
此外,研究还克隆并表征了一种新的尼古丁氧化还原酶基因(nicA2)。nicA2与之前已知的nicA1基因(编码尼古丁氧化还原酶)具有低氨基酸同源性(10.9%)。通过基因敲除实验,发现删除nicA2而非nicA1会阻止假单胞菌对尼古丁的降解,表明nicA2在尼古丁降解中起关键作用。本研究还构建了多个基因敲除突变株,包括mfs、sapd、pnao和nicA2基因敲除株,进一步证实了这些基因在尼古丁降解中的重要性。
通过蛋白质组学和转录组学的结合分析,本研究不仅揭示了尼古丁降解的新途径,还为环境污染物的生物降解提供了新的靶点。这些发现对于理解假单胞菌如何适应和降解环境中的有毒化合物具有重要意义,并可能为未来的生物修复和生物催化应用提供理论基础。
Bioscreen全自动生长曲线分析仪的应用
Bioscreen全自动生长曲线分析仪用于监测和比较不同菌株在特定培养条件下的生长情况。使用了野生型菌株Pseudomonas putida S16和多个基因敲除突变株(如S16dspm、S16dmfs、S16dsapd、S16dpnao和S16dnicA2)。这些菌株在含有尼古丁作为唯一碳源和氮源的培养基中进行培养。通过连续监测培养基中的光密度(OD)变化来记录菌株的生长曲线。Bioscreen每隔一定时间(如每小时)测量一次培养基的OD值,从而生成详细的生长曲线。通过Bioscreen C获得的生长曲线,研究人员能够直观地比较野生型菌株和各个基因敲除突变株在尼古丁培养基中的生长差异。
实验结果
通过蛋白质组学和分子遗传学方法,系统地揭示了假单胞菌(Pseudomonas putida S16)中尼古丁降解的未知途径。在尼古丁和甘油培养条件下,假单胞菌S16的蛋白质组包含1292种蛋白质,其中126种蛋白质表达差异显著。这些差异表达的蛋白质主要涉及运输、解毒、氨基酸代谢等功能,表明假单胞菌在尼古丁存在时会调整其蛋白质表达以适应环境压力。研究发现3-琥珀酰吡啶(SP)转化为6-羟基-3-琥珀酰吡啶(HSP)的关键步骤由一种多酶反应催化,涉及钼蝶呤结合氧化酶(spmA)、钼蝶呤脱氢酶(spmB)和(2Fe-2S)结合铁氧还蛋白(spmC),并以钼蝶呤胞嘧啶二核苷酸为辅因子。这一发现填补了尼古丁降解途径中的关键空白。克隆并表征了一种新的尼古丁氧化还原酶基因(nicA2),其编码的酶将尼古丁转化为N-甲基假胆碱。nicA2与已知的nicA1基因(编码尼古丁氧化还原酶)具有低氨基酸同源性(10.9%)。通过基因敲除实验,发现删除nicA2而非nicA1会阻止假单胞菌对尼古丁的降解,表明nicA2在尼古丁降解中起关键作用。构建了多个基因敲除突变株,包括mfs、sapd、pnao和nicA2基因敲除株,进一步证实了这些基因在尼古丁降解中的重要性。
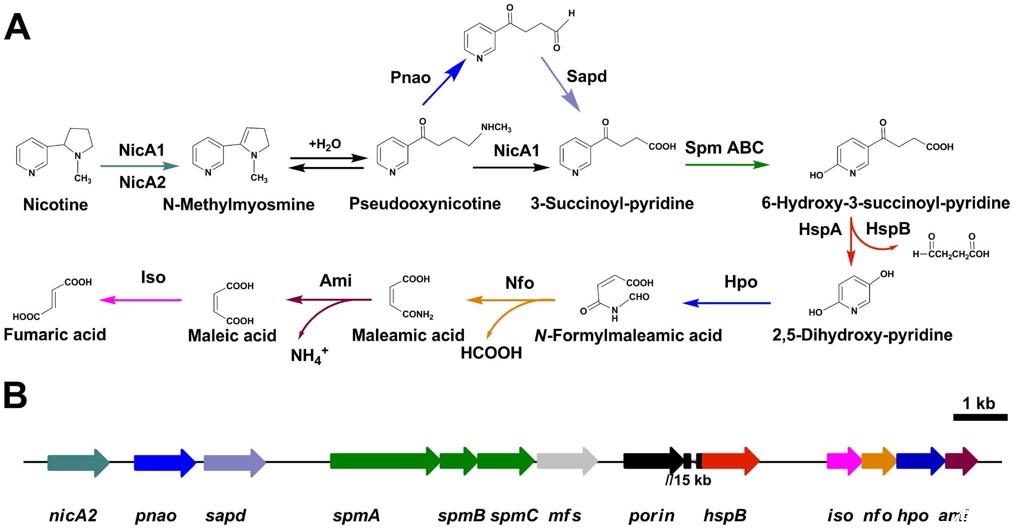
图1、铜绿假单胞菌S16的吡咯烷途径尼古丁代谢图。A.展示了S16中尼古丁分解代谢的完整步骤。
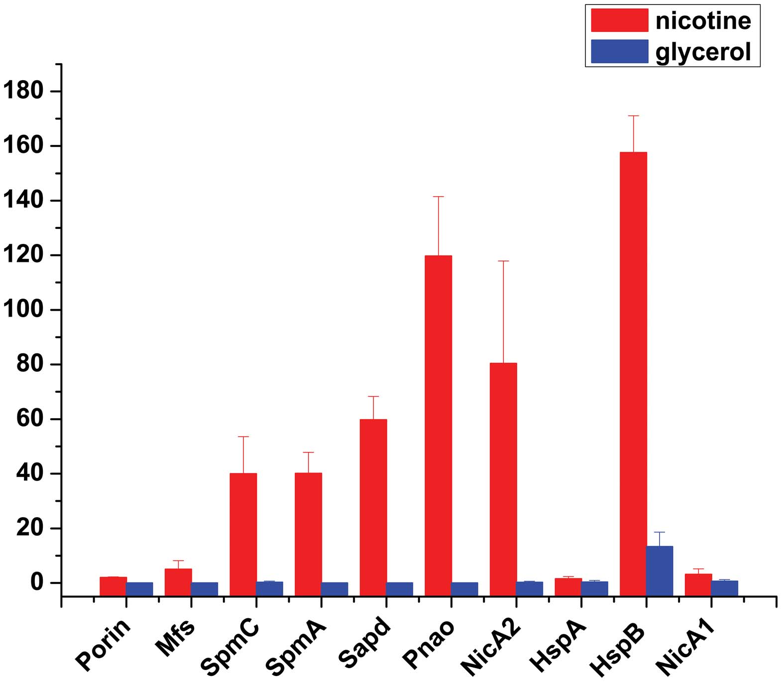
图2、尼古丁降解相关蛋白的表达水平变化。MS光谱计数检测到Porin、Mfs、SpmC、SpmA、Sapd、Pnao、NicA2、HspA、HspB和NicA1的表达。红色表示尼古丁培养基,蓝色表示甘油培养基。柱状图表示归一化光谱计数的平均值±标准误。
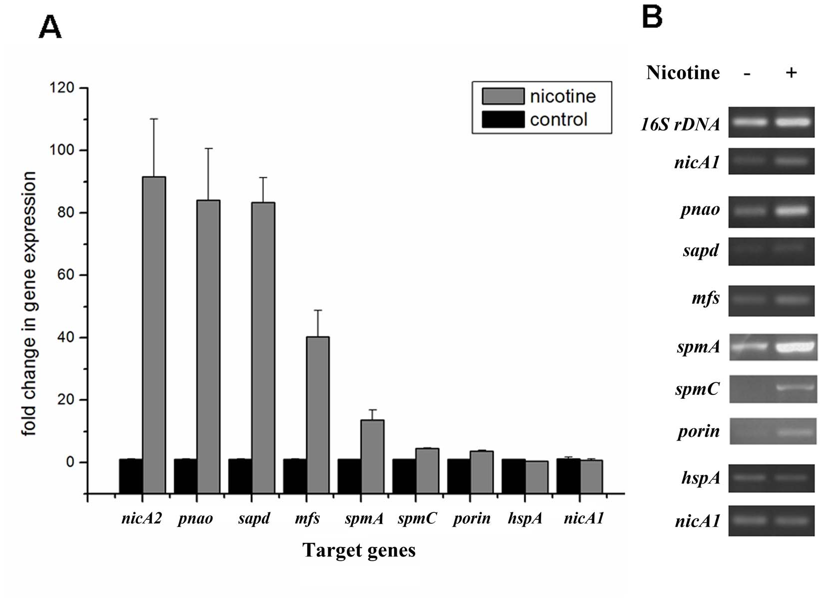
图3、差异表达蛋白的转录水平验证。A.RT-qPCR检测9个目标基因的mRNA表达水平,使用16S rRNA基因作为内参。结果为三次独立实验的均值±标准差。B.半定量RT-PCR验证目标基因转录情况,16S rDNA作为内控。
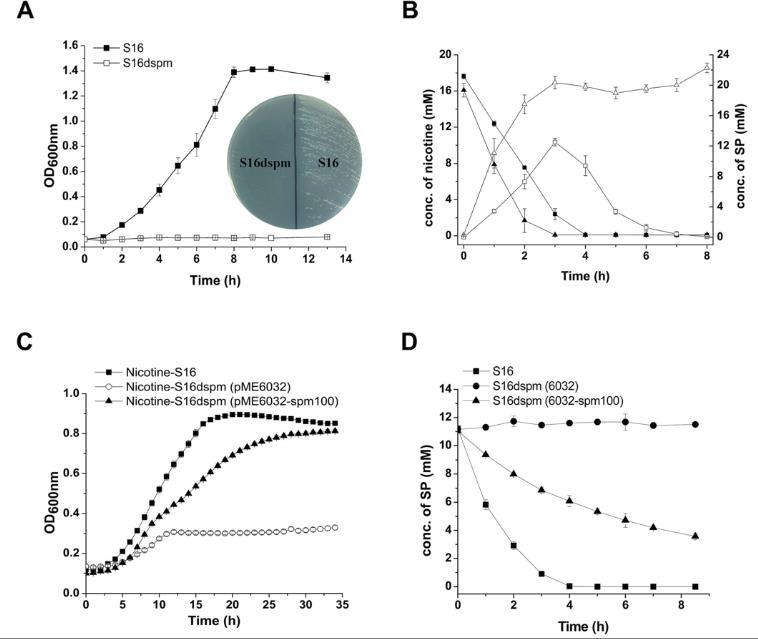
图4、spmABC基因的缺失与互补实验。A.野生型S16与spmABC缺失突变株S16Δspm在尼古丁培养基中的生长曲线。B.HPLC检测野生型与突变株对尼古丁的降解及SP的积累情况。C.野生型、突变株及携带互补质粒pME6032-spm100的菌株在尼古丁培养基中的生长曲线。D.HPLC检测野生型与不同质粒携带株对SP的降解情况。
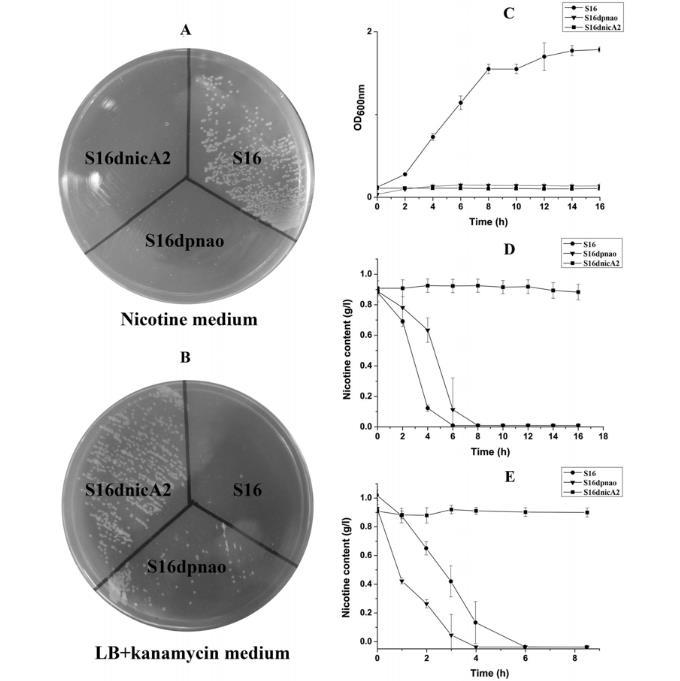
图5、野生型S16与基因缺失突变株的生长及静止态反应。A)在尼古丁平板上生长情况。B)在含卡那霉素的LB平板上生长情况。C)野生型、S16Δpnao、S16ΔnicA2在尼古丁培养基中的生长曲线。D、E)HPLC检测培养细胞与静止细胞对尼古丁的降解情况。
总结
铜绿假单胞菌(Pseudomonas putida)菌株是少数能够利用有毒和外源性化合物(如尼古丁)作为生长底物的微生物之一。虽然早在50多年前人们就发现了假单胞菌的尼古丁降解能力,但其潜在的分子机制一直不清楚。过去几年中,通过基因组文库筛选和野生型酶的纯化,做出了大量努力来鉴定3-琥珀酰吡啶(SP)羟基化的关键基因,但这些尝试均未能找到与SP羟基化相关的基因。
在本研究中,通过比较遗传学分析,本研究首次从P.putida S16中鉴定出三个关键基因spmA、spmB和spmC。由这些基因编码的异源三聚体酶需要钼蝶呤-胞苷二核苷酸(Mo-MCD)作为辅因子。以尼古丁或甘油为唯一碳源培养的S16蛋白质组中共检测到1292种蛋白质,为阐明尼古丁降解相关酶提供了详尽的信息。
通过比较尼古丁培养与甘油培养的细胞蛋白质组,研究人员发现了与尼古丁分解代谢相关的多种细胞过程和功能。Bioscreen C在本研究中发挥了关键作用,为研究者提供了可靠的生长分析数据,支持了基因功能的验证和尼古丁降解途径的解析。本研究不仅揭示了尼古丁降解的新途径,还为理解假单胞菌如何适应和降解环境中的有毒化合物提供了新的视角。这些发现对于未来的生物修复和生物催化应用具有重要意义。通过多学科的方法,全面揭示了假单胞菌S16中尼古丁降解的分子机制,为环境污染物的生物降解提供了新的靶点和理论基础。