2 结果与分析
2.1 巴氏链球菌与肺炎链球菌纤维二糖利用基因簇的同源性比对
如图1所示:以肺炎链球菌TIGR4两个纤维二糖相关的基因簇为参考,在猪源巴氏链球菌WUSP067中也存在2个潜在的纤维二糖相关的基因簇,将这2个基因簇分别命名为CL1和CL2,即E8M05_RS02040—E8M05_RS02055和E8M05_RS06225—E8M05_RS06270。CL1和CL2都包含编码磷酸转移酶系统(phosphotransferase systems,PTS)中的酶Ⅱ复合物和β-葡萄糖苷酶的基因。如图1-A,CL1中包含4个基因,分别是LicTwp(编码抗终止子蛋白)、BglP1wp(编码PTS中酶ⅡC)、BglP2wp(编码PTS中酶ⅡA)、BglAwp(编码6-磷酸-β-葡萄糖苷酶)。CL2包含10个基因,其中有5个基因据报道与纤维二糖利用相关。如图1-B所示,CL2包括celRwp(编码转录调控因子)、celAwp(编码6-磷酸-β-葡萄糖苷酶),以及celBwp、celCwp、celDwp(分别编码PTS中酶ⅡC、酶ⅡB、酶ⅡA),其余5个基因功能未知。在肺炎链球菌中,酶ⅡC特异性识别和捕获纤维二糖,6-磷酸-β-葡萄糖苷酶则负责将纤维二糖水解为单糖供细菌利用。
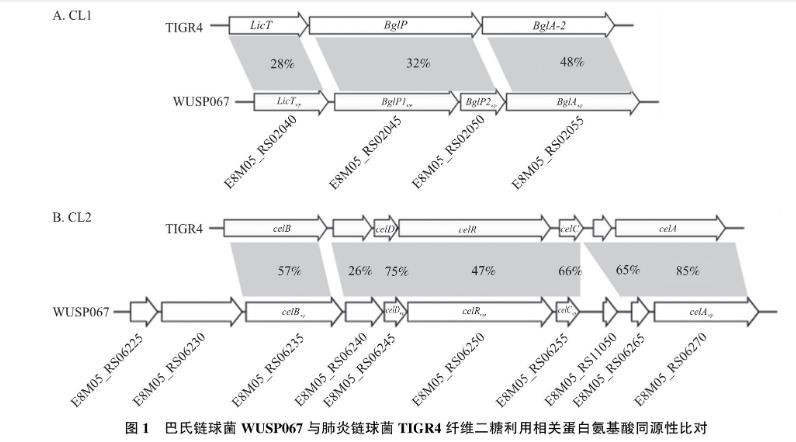
图1 巴氏链球菌WUSP067与肺炎链球菌TIGR4纤维二糖利用相关蛋白氨基酸同源性比对
2.2 纤维二糖对巴氏链球菌纤维二糖利用相关基因表达的影响
从图2可知,以添加10 g·L-1 Glu的DMEM培养基为对照组,在添加10 g·L-1 Cel的DMEM培养基中,celBwp和celAwp的基因表达量分别上调33倍和49倍(P<0.000 1),而Bglp1wp和BglAwp表达量无显著变化。此结果表明CL2在纤维二糖转运和利用中发挥作用,故选择CL2进行深入研究。
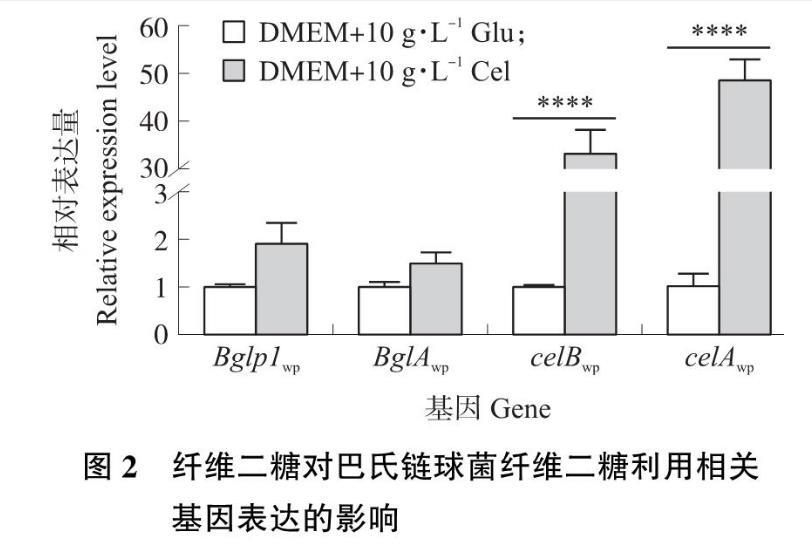
图2 纤维二糖对巴氏链球菌纤维二糖利用相关基因表达的影响
2.3 巴氏链球菌纤维二糖利用基因簇CL2结构
使用引物1-1-F/R、1-2-F/R、1-3-F/R、1-4-F/R扩增出长度分别为451、494、1 280 和350 bp的片段(图3-A、B); 使用引物1-6-F/R和1-7-F/R扩增出长度分别为501和439 bp的片段,而使用引物1-5F/R和1-8-F/R不能扩增出相应的条带(图3-C)。这表明CL2由2个操纵子组成,即E8M05_RS06225—E8M05_RS06255和E8M05_RS11050—E8M05_RS06270。
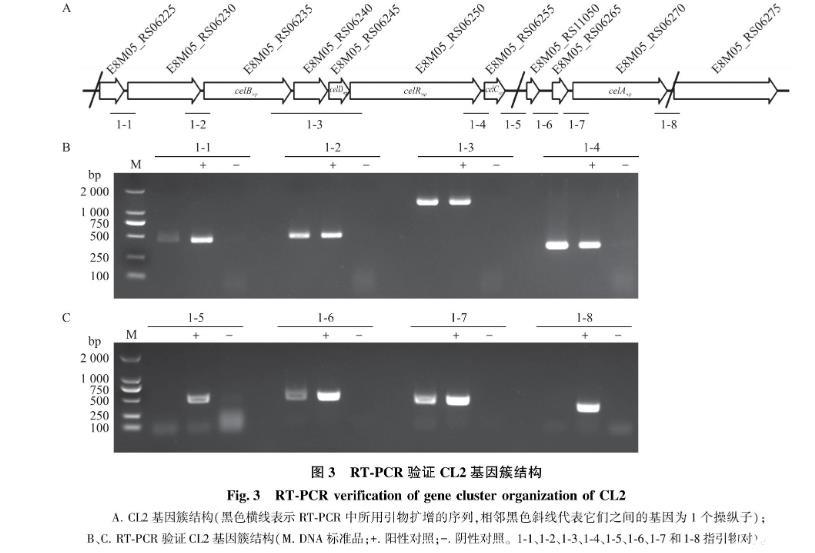
图3 RT-PCR验证CL2基因簇结构
2.4 缺失株ΔcelAwp的构建和鉴定
采用信号肽同源重组的方法构建ΔcelAwp(图4-A)。使用引物CelA-M/N,以缺失株ΔcelAwp为模板,扩增出大小为1 262 bp片段,而以野生株为模板,扩增出大小为1 928 bp片段; 使用引物CelA-X/Y,以缺失株ΔcelAwp为模板时,不能扩增出相应片段,而以野生株为模板时,可以扩增出大小为747 bp的片段(图4-B)。上述结果表明缺失株ΔcelAwp构建成功。
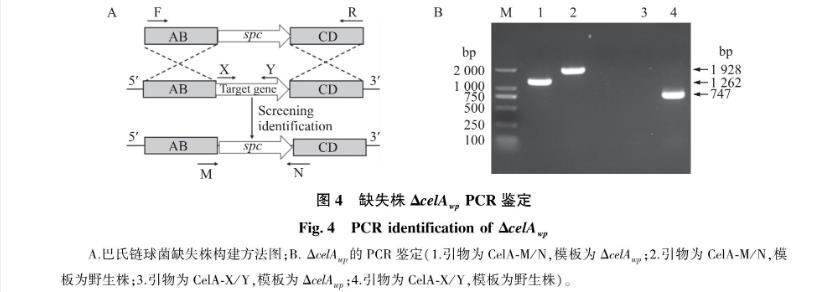
图4 缺失株ΔcelAwp PCR鉴定
2.5 celAwp对巴氏链球菌WUSP067生长的影响
如图5-A所示,在10 mL THB液体培养基中,将WUSP067野生株和缺失株ΔcelAwp在37 ℃、180 r·min-1摇床中培养,每隔2 h测定D600值,结果表明缺失基因celAwp对WUSP067在THB中生长没有影响。
如图5-B和5-C所示:在以葡萄糖为唯一碳源的培养条件下,缺失株生长情况与野生株一致; 在纤维二糖为唯一碳源的条件下,野生株能够生长,而ΔcelAwp则表现出明显的生长缺陷,生长12 h时D600低于0.5。以上结果表明,巴氏链球菌WUSP067能以纤维二糖为唯一碳源生长,celAwp基因参与纤维二糖的利用,缺失该基因影响巴氏链球菌对纤维二糖的吸收与利用。
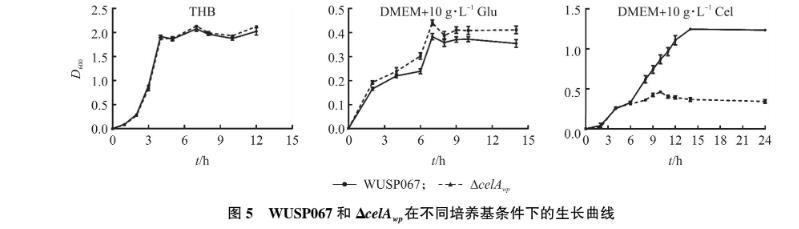
图5 WUSP067和ΔcelAwp在不同培养基条件下的生长曲线
2.6 celAwp对巴氏链球菌WUSP067毒力的影响
以ICR断奶仔鼠为感染模型进行WUSP067野生株和缺失株ΔcelAwp的毒力试验。结果如图6所示,攻毒剂量为每鼠3×108 CFU,攻毒后野生株攻毒小鼠3日内全部死亡,缺失株ΔcelAwp攻毒小鼠死亡时间延后,14 d后存活率20%,与野生株相比差异显著(P<0.05)。此结果表明,celAwp基因影响巴氏链球菌WUSP067的致病性,缺失该基因其毒力减弱。
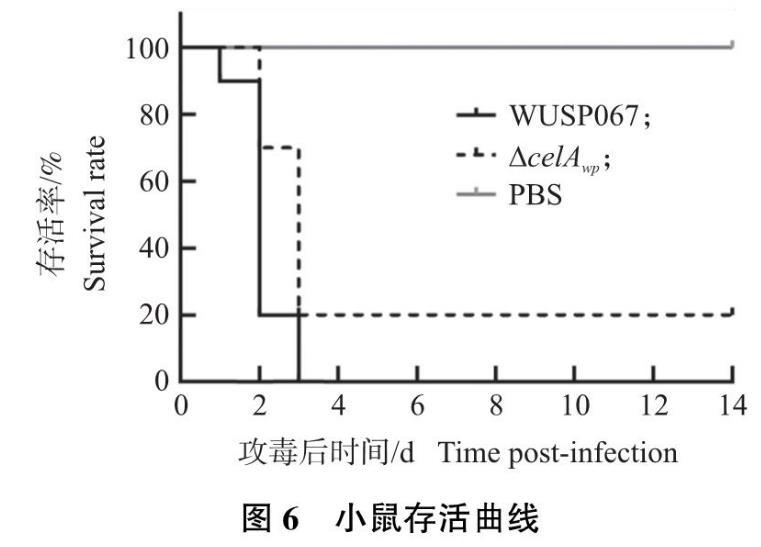
图6 小鼠存活曲线
3 讨论
目前,在链球菌属中,只有肺炎链球菌和变异链球菌中有纤维二糖利用基因簇的报道。在变异链球菌中仅有纤维二糖利用基因簇CL2,且未见其与致病性相关报道。猪源巴氏链球菌WUSP067中CL2与变异链球菌CL2各基因的覆盖率在90%以上,氨基酸同源性为56%~90%。当前,猪源巴氏链球菌全基因组序列仅有WUSP067(GenBank ID:NZ_CP039457.1)和WUSP074(GenBank ID:NZ_CP116958.1)2株菌,WUSP074也有纤维二糖利用基因簇CL1和CL2,其CL1与WUSP067的CL1覆盖率和氨基酸同源性均为100%,其CL2与WUSP067的CL2覆盖率90%,氨基酸同源性为56%~100%。
在肺炎链球菌中,纤维二糖转运和利用的基因簇主要有2个,都包含编码PTS中的酶Ⅱ复合物和6-磷酸-β葡萄糖苷酶的基因,前者负责纤维二糖的识别、磷酸化和转运,后者则将磷酸化后的纤维二糖水解为单糖供细胞利用。它们在调控方式上分为2种:一种是通过抗转录终止子蛋白调控,纤维二糖存在时被激活,抑制转录终止子的形成从而促进后续基因的表达; 另一种则是通过转录激活子celR调控,celR被纤维二糖激活后结合在纤维二糖利用操纵子的启动子区域,促进相关基因的表达,这种正调控方式也存在于变异链球菌和嗜热杆菌中,而艰难梭菌中celR是作为转录抑制子直接结合在纤维二糖操纵子的启动子区域来抑制基因的表达。本试验通过同源性分析发现巴氏链球菌WUSP067中预测存在2个潜在的纤维二糖利用基因簇CL1和CL2,预测其调控方式分别与第1种和第2种类似。从本研究中发现,CL1中BglP1wp、BglAwp基因不能感应纤维二糖,而CL2中的celBwp和celAwp基因能感应纤维二糖,表达量上调极显著,有助于巴氏链球菌WUSP067利用纤维二糖,表明CL2与纤维二糖利用相关。纤维二糖的利用与细菌的致病性相关:肺炎克雷伯菌中缺失celB基因(编码酶ⅡC)后在肠道定殖能力减弱; 肺炎链球菌中,缺失BglA3(编码β-葡萄糖苷酶)导致该菌对小鼠的致病性减弱; 艰难梭菌中,celA(编码酶ⅡB)突变株因无法在肠道中获取纤维二糖而表现出定殖劣势; 李斯特菌中,纤维二糖通过抑制毒力基因激活子PrfA来抑制相关毒力基因的表达。本试验结果表明,celAwp(编码6-磷酸-β-葡萄糖苷酶)缺失后,巴氏链球菌在纤维二糖为唯一碳源的培养条件下表现出明显生长缺陷,说明celAwp在纤维二糖的利用中发挥重要作用,随后通过小鼠致病性试验明确celAwp是巴氏链球菌WUSP067的毒力因子。
目前,由于尚无巴氏链球菌互补株构建方法,本试验仅构建了CL2基因簇中celAwp基因的缺失株,明确该基因与巴氏链球菌WUSP067纤维二糖的利用和其致病性相关,后续还应对celAwp编码的6-磷酸-β葡萄糖苷酶的酶活性进行测定,进一步明确其功能。巴氏链球菌可存在于猪的肠道中,故celAwp基因是否能通过利用纤维二糖促进巴氏链球菌在肠道中的竞争优势,帮助该菌在肠道中定殖,从而增强其致病性有待后续深入研究。此外,关于CL2的调控方式及该基因簇中其他基因的功能也值得进一步的探究。
综上,本试验结果明确猪源巴氏链球菌WUSP067能以纤维二糖为唯一碳源生长,CL2是纤维二糖利用相关基因簇,其中celAwp基因促进该菌利用纤维二糖,对该菌生长和毒力有显著影响,表明其在巴氏链球菌致病过程中发挥作用。