isp-1(qm150)发育迟缓的基因内抑制子也抑制寿命延长和线粒体未折叠蛋白反应的激活
我们确定了isp-1(qm150)突变体发育迟缓的抑制是否与寿命延长的抑制相关。携带发育抑制子等位基因的动物也减少了寿命延长,抑制程度取决于温度。在25°C时,isp-1(qm150sea5)和isp-1(qm150sea7)的寿命与N2相似,而isp-1(qm150sea4)寿命较短;而在15°C时,所有测试的sup突变都部分抑制了isp-1(qm150)的寿命延长(图2E)。
线粒体未折叠蛋白反应(mtUPR)在秀丽隐杆线虫中由线粒体应激诱导,通过hsp-6p::gfp报告基因的表达进行评估(18)。我们将hsp-6p::gfp品系与isp-1品系和三个抑制子品系进行杂交。所有三个isp-1(qm150)抑制子都降低了isp-1(qm150);hsp-6p::gfp品系中的hsp-6p::gfp荧光,表明mtUPR的激活也被减弱(图3 A和B)。
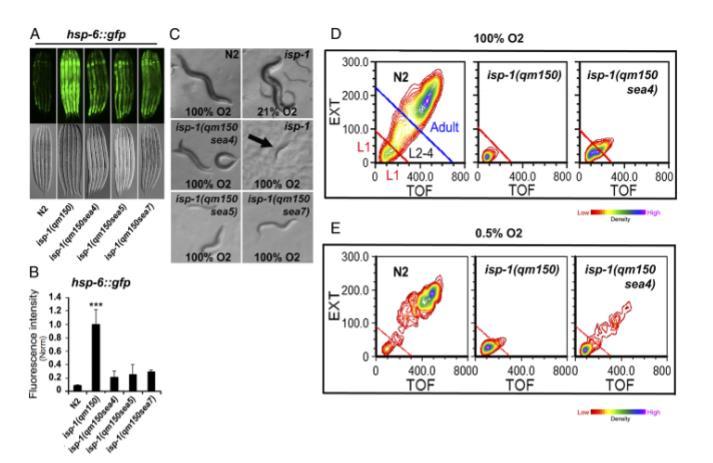
图3. 基因内抑制因子对isp-1(qm150)诱导线粒体未折叠蛋白反应、高氧敏感性和低氧敏感性的影响。(A) 抑制因子减弱hsp-6p::gfp表达(mtUPR反应)。表达hsp-6p::gfp线虫的落射荧光和DIC诺马尔斯基成像图。(B) 通过ImageJ软件(50)比较各品系hsp-6p::gfp荧光强度。(C) 基因内抑制因子部分挽救isp-1(qm150)的高氧致死性。N2在100% O2环境下可产生下一代;isp-1(qm150)在21% O2环境下可产生下一代;isp-1(qm150)在100% O2环境下于L1期死亡;基因内抑制因子在100% O2环境下可发育至L3/4期。图像为同步化L1幼虫在25°C接种至NGM-OP50培养基后第8天拍摄。(D和E) 高氧与低氧环境下经COPAS分选的线虫。N2线虫在25°C培养第2天分选,其他品系在第5天分选。请注意N2的L1幼虫来自其下一代。误差线:标亮实验的标准误,*P<0.05 **P<0.01 ***P<0.001。
isp-1(qm150)突变导致对高氧和低氧的敏感性,这种敏感性被系链区域的突变所抑制
暴露于高于或低于大气水平(21%)的氧气浓度会增加ROS的产生。正如先前报道的(21),我们观察到野生型动物的发育和繁殖不受高氧(100%氧气,90 kPa)的显著影响。相比之下,在高氧条件下观察到一个先前未报道的缺陷,即isp-1(qm150)表现出对高氧的深刻敏感性,未能发育,最终在3-5天后作为L1/2幼虫死亡。系链区域突变赋予了对这种表型的部分抑制,因为携带isp-1(qm150)抑制子的动物发育到L4阶段;然而,它们从未达到成年繁殖阶段(图3 C和D,表2和SI附录,图S2)。
此外,缺氧(1%氧气;参考文献22)也会加剧呼吸缺陷突变体(如isp-1(qm150))的ROS产生,我们观察到isp-1(qm150)动物在25°C的缺氧条件下停滞在L1/2发育阶段并死亡(图3E和SI附录,图S3)。在这种条件下,系链区域突变允许isp-1(qm150)动物进一步发育。
isp-1(qm150)系链中的突变增加线粒体功能并稳定呼吸体
为了直接评估基因内抑制子对氧化ATP产生能力的影响,我们测量了完整分离线粒体的状态3(ADP刺激)呼吸。我们比较了从N2、isp-1(qm150)和一个抑制子isp-1(qm150sea4)分离的线粒体的速率。线粒体是从在常氧、21±2°C下生长的第1天成虫中分离的。我们发现,当电子从苹果酸或琥珀酸底物提供给复合物III上游的呼吸链时,isp-1(qm150)线粒体中的状态3呼吸受损(图4 A-F),正如其他人报道的那样。相比之下,当电子通过抗坏血酸加N,N,N',N'-四甲基对苯二胺直接提供给细胞色素c(cyt c)(复合物III下游)时,呼吸不受影响(图4 C和D,红色星号表示在添加TMPD/抗坏血酸绕过复合物III之后)。
通过TMPD/抗坏血酸进行的isp-1(qm150)分离线粒体的完整呼吸证实了isp-1(qm150)在复合物III处存在电子传递缺陷。在isp-1(qm150sea4)中,状态3呼吸速率显著更高,接近野生型功能,表明复合物III得到挽救(图4 A-H和SI附录,图S4)。此外,据报道isp-1(qm150)通过破坏由复合物I、III和IV形成的呼吸体超复合物来影响复合物I依赖性呼吸。在蓝绿原生胶中,我们在来自所有N2和所有isp-1(qm150;sea4)样品的I,III和I,III,IV超复合物条带中观察到通常归因于复合物I存在的黄递酶活性,但在isp-1(qm150)样品中没有观察到(图4J)。我们还分析了三个抑制子(sea4、sea5和sea7)的CO2输出,并将它们的CO2输出与N2和isp-1(qm150)进行了比较。我们观察到isp-1(qm150)突变体产生的CO2显著低于N2,而isp-1(qm150sea4)突变体部分恢复了CO2输出(图4K)。
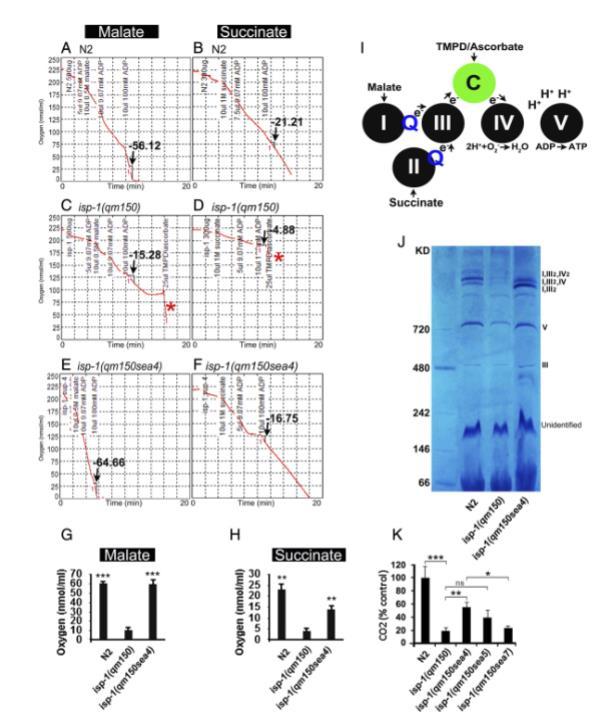
图4. 完整离体线粒体的ADP刺激呼吸作用。图中展示了N2、isp-1(qm150)和isp-1(qm150sea4)的氧耗曲线。(A) 使用苹果酸盐底物作为上游(复合物I)电子供体时,N2的ADP刺激状态3速率(箭头所示)。添加2 mM ADP的状态3呼吸作用代表N2线粒体氧化磷酸化的最大能力。(B) 使用琥珀酸盐底物作为上游(复合物II)电子供体时N2的氧耗曲线。(C-F) 使用复合物III上游底物(苹果酸盐和琥珀酸盐)时,isp-1(qm150)和isp-1(qm150sea4)的ADP刺激状态3速率(箭头所示)比较。在C和D中,星号(.)表示添加TMPD/抗坏血酸(细胞色素c的电子供体,位于复合物III下游)后isp-1(qm150)的氧耗量。(G和H) 展示了3个生物学重复的状态3测量定量结果。P值与isp-1(qm150)的三次重复实验进行比较。
H中P值:N2: 0.000204,isp-1(qm150sea4): 0.000707。I中P值:N2: 0.002405,isp-1(qm150sea4): 0.009303。完整离体线粒体的呼吸作用使用克拉克电极测量。呼吸速率的传统单位A0/min/mg蛋白相当于nmol 1/2O2/min/mg蛋白(52)。(f) 示意图描绘电子从电子供体底物(苹果酸盐、琥珀酸盐或TMPD/抗坏血酸)通过线粒体ETC复合物的流动。ADP刺激呼吸作用(状态3)是因为其通过复合物V磷酸化为ATP允许质子流经复合物I、III、IV(出)和V(入)。注意来自TMPD/抗坏血酸的电子流与功能性复合物III无关。
(J) 蓝色原生凝胶。复合物1存在于N2和isp-1(qm150sea4)的超复合物条带中,但不存在于isp-1(qm150)。(K) isp-1(qm150)的CO2输出与抑制株和N2比较(每个品系:n=1,000只COPAS分选的同步化年轻成年虫,体型相同(可能重量也相同),进行三次技术重复(每个品系/条件3000只虫)。误差线:突出显示实验的标准误,*P<0.05,**P<0.01,***P<0.001。
相关新闻推荐
1、3种植物精油对调理牛排腐败贮藏过程优势腐败菌生长抑制情况及细菌生长曲线(一)
3、针对内毒素及微生物,分享医疗器械质量管理体系构建及环节管控重点