1.2.7噬菌体紫外线敏感性测定
取噬菌体原液2 mL于直径9 cm无菌培养皿中,置于超净工作台紫外灯下30 cm照射,间隔5 min取样,采用双层琼脂平板法测定噬菌体效价,所有试验均重复3次。
1.2.8噬菌体裂解谱测定
测定菌株包括13株枯草芽孢杆菌、5株莫哈韦芽孢杆菌、1株蜡样芽孢杆菌及1株地衣芽孢杆菌,为实验室现有菌种库保存。复苏菌株后挑取单菌落接种于LB液体培养基中,37℃摇床振荡培养4 h至对数期,取100μL对数期菌液与50℃LB半固体培养基混合制备双层板,上层凝固后在双层平板中央滴加10μL噬菌体(108 PFU/mL)悬液。37℃温箱培养6 h观察是否出现噬菌斑。
1.2.9噬菌体体外裂解试验
将噬菌体富集液10倍比稀释,浓度依次为108 PFU/mL、107 PFU/mL、106 PFU/mL、105 PFU/mL、104 PFU/mL。分别取不同浓度的噬菌体稀释液100μL加入96孔板,再向每孔中加入100μL浓度为107 CFU/mL的宿主菌液,每个梯度做3个平行,以200μL菌液作为阳性对照,以200μL LB液作为阴性对照。使用动态酶标仪,测定其OD600值,每隔1 h测定一次,共测定24 h。
1.2.10抗性菌株筛选
挑取噬菌体裂解的菌株单菌落LB液体培养,培养至对数期后按最佳MOI比例加入噬菌体,37℃180 r/min培养至澄清后,取1 mL裂解液转接至对数期宿主菌菌液中共培养,如此重复多次直至培养液出现浑浊后,使用接种棒涂布于LB固体平板,37℃培养12 h,挑取单菌落使用点斑法进行裂解试验,直到筛选出噬菌体不能裂解且稳定的抗性菌株。
1.2.11抗性菌株生长动力学监测
将抗性菌株与原始菌株接种至LB液体培养基中,37℃、160 r/min振荡培养6 h作为种子液。将其接种至96孔板,100μL菌液与100μL LB液体培养基,使用酶标仪于37℃培养并测定OD600,每隔1 h测定一次,测定48次,设置3个重复。
1.2.12抗性菌株发酵效价测定
同1.2.11培养种子液,按2%接种量转接至新鲜LB培养基进行摇菌发酵,每隔12 h取样测定效价,测定4次。效价测定采用稀释涂布法,使用LB液进行梯度稀释,吸取菌液100μL涂布LB固体平板,每个稀释度设置3个重复,37℃倒置培养12 h后计数菌落形成单位(CFU/mL)。
1.2.13淀粉酶活性测定
采用DNS法测定α-淀粉酶活力,使用LB液体培养基进行菌株发酵,测定8 h、12 h、24 h、36 h、48 h淀粉酶活性。取发酵液10 000×g离心6 min,取上清液进行检测。使用α-淀粉酶试剂盒进行淀粉酶活性测定。使用试剂处理后,OD540读取光密度值并代入标准曲线方程进行计算。
2结果
2.1噬菌体PJNB028形态
枯草芽孢杆菌噬菌体命名为PJNB028。如图1所示,噬菌斑近圆形,边缘有晕环,直径约2 mm。
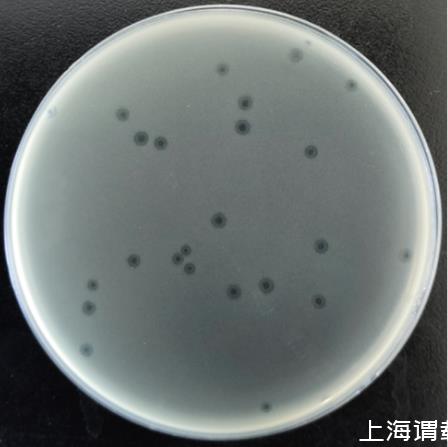
图1噬菌体PJNB028噬菌斑形态
2.2噬菌体PJNB028全基因组分析
噬菌体PJNB028基因组为双链环状DNA,全长162 308 bp,G+C含量为34%。全基因组BLASTN比对结果表明,噬菌体PJNB028与枯草芽孢杆菌噬菌体Bacillus phage 2S-4相似度最高为100%。经CARD网站预测PJNB028基因组中未发现耐药基因、毒力基因。全基因组序列已提交至国家微生物科学数据中心(National Microbiology Data Center,NMDC),编号NMDC60203838。
如图2所示,PJNB028具有237个开放阅读框(open reading frame,ORF),含有13个tRNA基因(表1),66个ORF编码为已知功能蛋白质。其中已知功能阅读框分为5个功能模块:裂解、复制、结构与包装、转录调控和未知模块。2个ORF与裂解相关,30个ORF与DNA复制修复相关,18个ORF是结构与包装蛋白,16个ORF与转录调控相关,其余为假定蛋白(hypothetical protein)。在ORF160和ORF161之间编码了tRNA-Arg、tRNA-Arg、tRNA-Gly、tRNA-Ser、tRNA-Ser、tRNA-Thr、tRNA-Leu、tRNA-Cys和tRNA-Met,在ORF161和ORF162之间编码了tRNA-Pro、tRNA-Glu、tRNA-Gly和tRNA-Asn。
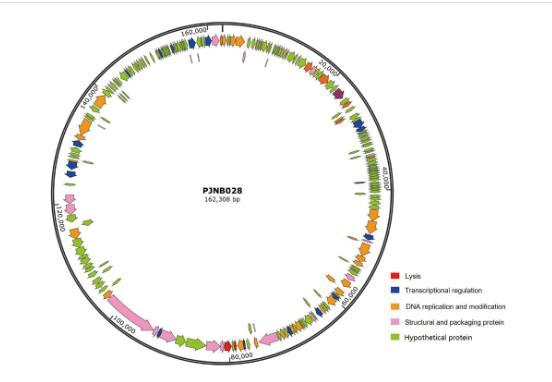
图2噬菌体PJNB028基因组圈图
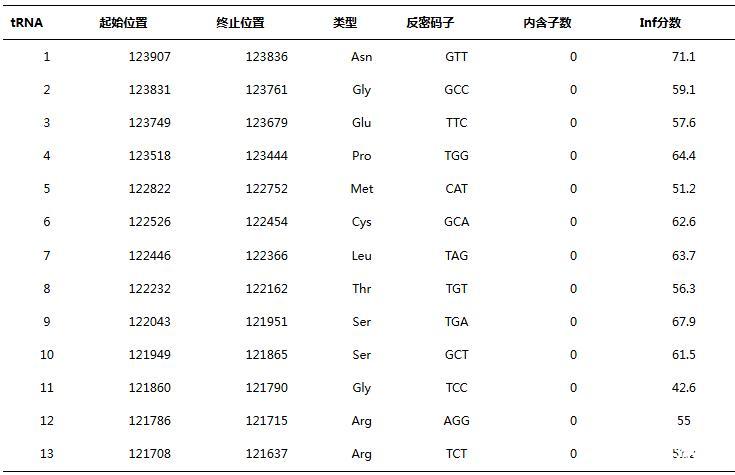
表1噬菌体PJNB028基因组中的tRNA基因
其中,与裂解相关的ORF131为穿孔素,作用于细菌细胞膜,是一种疏水性跨膜蛋白。ORF133预测为LysM肽聚糖结合域蛋白,可能与细菌细胞壁的识别和相互作用有关,两者相互配合联合抑菌。18个ORF与蛋白质的结构和包装有关,ORF138(phage tail spike protein)结合多糖特定细菌宿主,有助于决定噬菌体侵染的专一性;ORF140(phage tail domain-containing protein)为噬菌体的尾部结构;ORF141(phage tail tape measure protein)用于测量噬菌体尾部长度,控制尾部长度,有助于噬菌体识别和结合宿主细菌。ORF157预测为末端酶大亚基,末端酶对启动DNA包装至关重要。30个ORF与DNA复制相关,PJNB028编码了许多与核酸修饰有关的蛋白质,如ORF86DNA拓扑异构酶、ORF104DNA连接酶、ORF128交叉连接内脱氧核糖核酸酶RuvC等。通过NCBI结构域预测发现ORF86是四型拓扑异构酶。ORF33预测为含有DNA样解旋酶C端结构域的蛋白质,与DNA的复制和修复有关。ORF142(site-specific integrase)是一种位点特异性重组酶,能够在特定DNA序列之间进行重组。16个ORF与转录调控相关。ORF175(helix-turn-helix domain-containing protein)是含有螺旋-转角-螺旋结构域的蛋白质,有研究表明该蛋白质可用于调控CRISPR系统。ORF202(Rho termination factor N-terminal domain-containing protein)主要与转录终止调控相关。ORF221(type II toxin-antitoxin system HicB family antitoxin)II型毒素-抗毒素系统是一种广泛存在于细菌和古菌中的遗传系统,有利于噬菌体PJNB028在细菌细胞内生存。
相关新闻推荐
1、流感重配病毒以不同MOI感染MDCK细胞的生长曲线(一)